



原标题:QoI类杀菌剂应用与抗性机制研究进展
1 甲氧基丙烯酸酯类杀菌剂概况
甲氧基丙烯酸酯类(QoIs)杀菌剂是继苯并咪唑类、三唑类后第3个里程碑式的杀菌剂类别,被广泛用于防治作物病害。该类杀菌剂具有广谱、渗透、快速分布等特点和较好的环境相容性,可用于茎叶喷雾、种子处理,也可进行土壤处理。其在植物体内的传导活性较强,不仅能阻止真菌在植物体内侵入和扩展,而且能明显地降低初侵染和再侵染的孢子基数以达到防治病害的目的。该类杀菌剂具有良好的内吸性和熏蒸作用,被叶片吸收后可在木质部中随水流在运输系统中流动(即内吸性),此外,其可在叶片表面的气相中流动,当其随着气相进入叶片后又可在木质部中流动(即熏蒸作用),使得活性成分能够有效再分配和充分传递。甲氧基丙烯酸酯类杀菌剂耐雨水冲刷性能好、持效期长,因此,其活性不受应用环境影响,在晴雨天气均能提供良好的保护和治疗作用。该类杀菌剂对卵菌纲、藻菌纲、子囊菌纲和半知菌纲病害,如白粉病、锈病、颖枯病、网斑病、霜霉病、稻瘟病等均有良好的活性,且使用剂量低,适用于禾谷类、果树和蔬菜等多种作物。在我国,QoI类杀菌剂在各类粮食作物和经济作物病害防治上均有登记。
1.1 作用机理
国际杀菌剂抗性委员会(FRAC)基于交互抗性模式和作用方式将甲氧基丙烯酸酯类杀菌剂归类于C3(Group 11)(图1)。该类杀菌剂是以天然源物质为基础,经结构修饰和化学优化而具有农药活性,主要作用于真菌线粒体,与线粒体上细胞色素b的Qo位点结合,阻断细胞色素b和细胞色素c1之间的电子传递,阻止NADH的氧化和ATP的合成,抑制真菌呼吸作用、破坏其内部的能量循环从而抑制真菌生长(图2)。
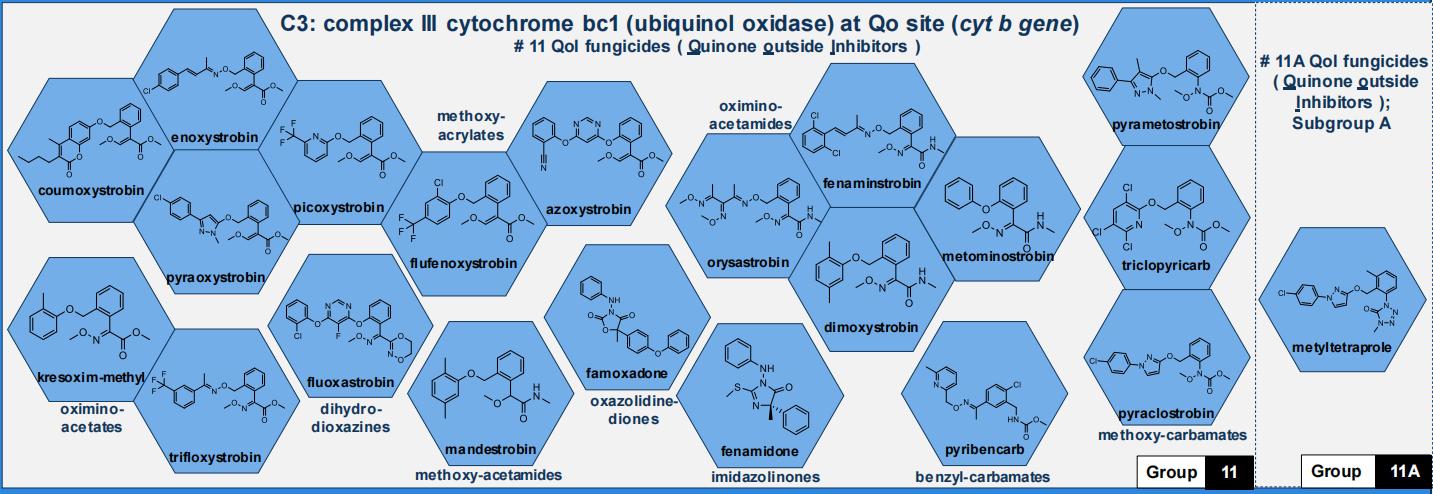
图1 基于交互抗性模式和作用方式的杀菌剂划分
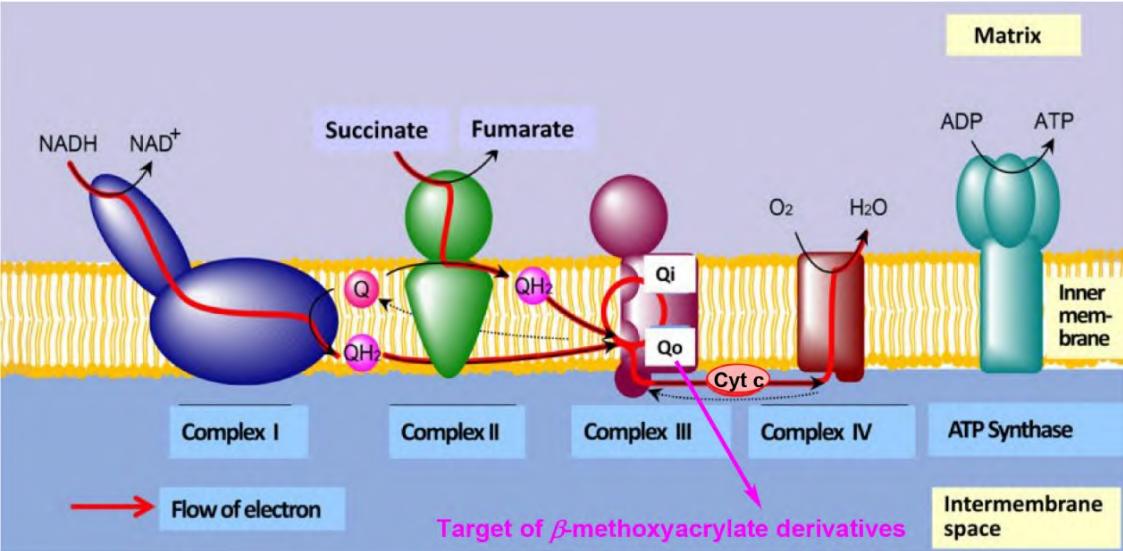
图2 甲氧基丙烯酸酯衍生物靶标
甲氧基丙烯酸酯类杀菌剂与目前其他类型的杀菌剂不存在交互抗性关系,但与其同类不同活性基团的杀菌剂之间(FRAC code 11)存在交互抗性,因此,该类杀菌剂对防治已对吗啉类(morpholines)、三唑类(triazoles)、二羧酰胺类(dicarboximides)、苯基酰胺类(phenylamides)、琥珀酸脱氢酶抑制剂类(SDHIs)等杀菌剂产生抗性的植物病原菌菌株非常有效。
1.2 种类及应用
目前已开发的甲氧基丙烯酸酯类杀菌剂多达20余种,广泛应用于草坪、蔬菜、水果、谷类、核果类等植物。各品种甲氧基丙烯酸酯类杀菌剂的主要化合物结构存在着一定的差异,因而其内吸传导性、生物活性、防治对象及适用作物存在不同。目前商品化的主要品种包括嘧菌酯、吡唑醚菌酯、肟菌酯、氟嘧菌酯、啶氧菌酯等。
嘧菌酯(azoxystrobin)由先正达开发并于1997年上市,1999年获中国行政保护。根据作物和病害的不同该杀菌剂施用剂量为25~400 g a.i./hm2,通常使用剂量为100~375 g a.i./hm2。如在25 g a.i./100 L剂量下,对葡萄霜霉病有很好的预防作用;在12.5 g a.i./100 L剂量下,对葡萄白粉病和苹果黑星病有很好的防治效果;在200 g a.i./hm2剂量下,对马铃薯疫病具有预防作用。嘧菌酯是先正达杀菌剂产品线的重要组成部分,一度占据全球杀菌剂年销售额榜首,年销售峰值超过10亿美元。2019年,该杀菌剂全球销售额14.89亿美元,2014—2019年复合年增长率为4.15%,近年来占据的全球市场份额始终保持在前10位。
吡唑醚菌酯(pyraclostrobin)由巴斯夫公司2000年研发,并于2002年上市。该杀菌剂应用范围较广,现已在全球100个多国家登记,用于100多种作物,其中在我国登记的作物有80余种。研究表明,吡唑醚菌酯对多种病原菌有效,例如谷物叶枯病菌(Septoria tritici)和条纹病菌(Pyrenophora teres),大豆褐纹病菌(Septoria glycines)、紫斑病菌(Cercospora kikuchii)和锈病病菌(Phakopsora pachyrhizi),荔枝霜疫霉菌(Peronophythora litchii),葡萄白粉病菌(Uncinula necator),番石榴叶斑病菌(Pestalotiopsis disseminatum)等。吡唑醚菌酯在食用作物上的用量为50~250 g a.i./hm2,草坪上的用量为280~560 g a.i./hm2。巴斯夫已在我国推出18个基于吡唑醚菌酯的高效杀菌和对植物具有保健作用的施乐健®系列产品,包括百泰、凯润、健达等,这些产品对几乎所有作物具有病害防治作用及植物保健作用。同时,吡唑醚菌酯作为“有病治病,无病健体”理念的典范,是美国环保署、欧盟和中国就“植物健康作用”登记的第一个产品。2019年吡唑醚菌酯全球销售额10.15亿美元,2014—2019年复合年增长率为0.30%。
肟菌酯(trifloxystrobin)于1999年在南非和瑞士上市,被认为是第2代甲氧基丙烯酸酯类杀菌剂,对白粉病、叶斑病有特效。大豆是肟菌酯的第一大用药作物,2015年,大豆用肟菌酯的销售额为3.10亿美元,占其全球市场的47.7%;2016年,大豆用肟菌酯销售额为3.03亿美元,占其全球市场的47.3%。自上市以来,肟菌酯全球市场总体呈增长趋势。2015年,由于全球大豆病害抗性的加剧、气候条件不利以及转基因作物的大量种植等多方面因素,导致肟菌酯销售额略有下降,但在2018年农化市场逐渐复苏,2019年肟菌酯全球销售额为8.42亿美元,2014—2019年复合年增长率为4.68%。肟菌酯适配性强,具有较多混配产品,三唑类杀菌剂是其最好的配伍类型。肟菌酯可与三唑类杀菌剂三唑醇、苯醚甲环唑、丙环唑、环丙唑醇、戊唑醇、三唑酮等进行混配,还可与琥珀酸脱氢酶抑制剂类杀菌剂氟唑菌苯胺、氟吡菌酰胺、啶酰菌胺等进行混配。
氟嘧菌酯(fluoxastrobin)由拜耳公司于1994年发现并于2004年在欧洲首先上市。作为全新一代QoI类杀菌剂,具有优于嘧菌酯、肟菌酯和吡唑醚菌酯等同类产品的内吸活性、跨层传导和木质部传导性。该杀菌剂可有效防治100余种真菌病害,对种传和土传病害腥黑穗病、黑穗病等,小麦叶部病害均具有优异防效。田间药效试验结果表明,氟嘧菌酯对小麦叶枯病、颖枯病和锈病,大麦云纹病、条纹病和锈病等的防效均达到或超过醚菌酯,但对大麦白粉病或网斑病等气传病害的防效较差。氟嘧菌酯使用量一般为75~200 g a.i./hm2,用作禾谷类作物的种子处理剂时,用量为5~l0 g a.i./100 kg。由于该杀菌剂制剂产品可与众多产品配伍,且目前专利到期,国内多家企业已启动该产品的登记和上市工作,并获批开展氟嘧菌酯单剂和复配制剂的田间试验,因此,氟嘧菌酯在未来存在着巨大的市场潜力,很可能成为继嘧菌酯后又一优良产品。2018年,氟嘧菌酯全球销售额为2.45亿美元,2013—2018年的复合年增长率为3.9%。
啶氧菌酯(picoxystrobin)由先正达2001年在欧洲首次推出,2006年由杜邦公司收购后,先后在拉美、北美市场登记,在我国于2012年7月获得临时登记并于同年11月获正式登记。啶氧菌酯是甲氧基丙烯酸酯类杀菌剂中内吸活性最强的杀菌剂,与嘧菌酯和吡唑醚菌酯等比较,对小麦叶枯病、网斑病和云纹病有更强的治疗效果。据报道,该药剂比嘧菌酯和肟菌酯等具有更好的治疗活性,且啶氧菌酯在小宗作物上的表现丝毫不输其同类药剂,在我国的主要登记也是集中在小宗作物上用来替代嘧菌酯。使用啶氧菌酯防治谷物病害时,谷物的产量和质量明显提高,在生长期无病害发生,绿叶始终保持完好。冬小麦大田试验表明,用啶氧菌酯处理的小麦比用肟菌酯处理的平均增产0.2 t/hm2。在欧洲小麦试验田进行的试验中,用啶氧菌酯处理过的小麦产量与对照相比增产22%,比使用醚菌酯和三唑类杀菌剂氟环唑(epoxiconazole)混剂处理的增产0.4 t/hm2,这表明啶氧菌酯在小麦病害防治中效果好且增产显著。2006年,杜邦公司收购啶氧菌酯后,通过一系列新增登记以及开发复配产品等方式来提升销售额,其与环丙唑醇和苯并烯氟菌唑的复配产品热销于拉美地区,用于防治大豆、谷物和玉米病害。2019年,随着新复配产品的增多,啶氧菌酯的全球销售额上升至4.85亿美元,2014—2019年复合年增长率为6.74%。2018年起啶氧菌酯不再在欧洲国家继续登记,但这并未影响其在全球其他地区的销售,其在国内乃至整个亚洲的市场份额还在不断上升,国内厂家正在进一步挖掘啶氧菌酯的市场潜能,啶氧菌酯的开发和利用仍处于上升趋势。
2 甲氧基丙烯酸酯类杀菌剂抗性发生现状
甲氧基丙烯酸酯类杀菌剂作用位点单一,属于高抗药性风险类杀菌剂,在实际应用过程中极易产生抗药性。随着生产上频繁使用和用药量不断上升,导致各类病原菌抗性问题日趋严重,给全球粮食安全问题提出了严峻考验。
2.1 子囊真菌对QoI类杀菌剂抗性发生现状
在QoI类杀菌剂上市仅2年后,1998年首次在德国的谷物类作物中发现小麦白粉病菌(Blumeria graminis)的QoI类杀菌剂耐药菌株,随着全球该类杀菌剂的用量增加和使用范围扩大,抗性报道逐渐增多。Sierotzki等2000年在欧洲小麦白粉病菌中发现抗性菌株,而在之后的十几年间,小麦白粉病菌的抗药性在世界范围内仍在不断蔓延。2008年Sedláková等报道,德国、法国、比利时、丹麦和英国均发现了小麦白粉病菌对肟菌酯不同程度的抗性,在德国最高抗性菌株分布达到70%~90%。2017年,在澳大利亚的小麦田中同样也发现了小麦白粉病菌抗性菌株。2021年,Cowger等在美国中部和东部15个州共分离出381株小麦白粉病菌,它们对吡唑醚菌酯敏感度适中,RF最大值为11.2(抗性因子RF指杀菌剂对目标菌株与敏感菌株的EC50之比)。
以小麦白粉病为代表的气传病害传播速度较快,目前,在其他白粉病中也陆续检测到QoIs抗性。1999年,美国纽约256株葡萄白粉病菌(Erysiphe necator)中有2%能够在嘧菌酯2 mg/L的剂量下存活。Wong等2002年研究发现QoI类杀菌剂对美国葡萄白粉病菌存在不同程度的抗性,其中肟菌酯抗性最高(EC50:0.015 mg/L),其余依次为嘧菌酯(EC50:0.013 mg/L)、吡唑醚菌酯(EC50:0.00442 mg/L)。仅过了10年,Miles等于2012年在对美国密歇根州的12株葡萄白粉病菌株的研究中发现1株抗性突变体,其能在添加100 mg/L肟菌酯的培养基上正常生长,该菌株对肟菌酯的抗药性是Wong等在2002年所报道抗性的6,667倍。当前,世界各地均有不同程度的葡萄白粉病菌对QoI类杀菌剂的抗药性发生,在欧洲已有多国出现QoIs抗性报道。Hall等2016年测得吡唑醚菌酯对澳大利亚72个葡萄白粉病菌株的EC50值范围为0.0005~14.4 mg/L,其中42%的菌株EC50值在1 mg/L以上。2008—2013年采集自欧洲和澳大利亚的239株苹果白粉病菌(Podosphaera leucotricha)对吡唑醚菌酯较敏感,EC50值范围为1.39~2.07 mg/L,没有发现抗药性菌株。Heick等2015—2018年在丹麦和瑞典采集的43株甜菜白粉病菌(Erysiphe betae)中监测到25个对吡唑醚菌酯的抗性菌株。
Frederick等研究发现,纽约和美国东部的果园苹果黑星病菌(Venturia inaequalis)对QoI类杀菌剂抗性较低,67%的供试菌株对QoI类杀菌剂肟菌酯表现为敏感,仅有28%表现为抗性。直到2020年,Jaklová等从捷克多地采集的苹果黑星病菌中发现,抗性菌株多达95%。2021年,Turan等在意大利北方对49个苹果黑星病菌样品进行敏感性测定,结果显示,70%以上菌株对肟菌酯表现出抗药性,EC50值大于0.12 mg/L。由此可见,全球范围内苹果黑星病菌对QoI类杀菌剂的抗性水平在迅速升高。
2.2 半知真菌对QoI类杀菌剂抗性发生现状
2013年,日本采集的376个甜菜褐斑病菌(Cercospora beticola)中有15个对嘧菌酯和醚菌酯都有抗性。2015年,Piszczek等从波兰采集的甜菜褐斑病供试菌株中,有56%在添加1 mg/L嘧菌酯的PDA培养基中能正常生长,抗性株最高EC50值大于100 mg/L。2018年,Zakariae等从摩洛哥采集的甜菜褐斑病供试菌株中至少70%的菌株对嘧菌酯具有抗性,EC50值大于10 mg/L。日本和波兰甜菜褐斑病菌抗性均为首次报道。段亚冰等在温室中采集到的黄瓜褐斑病菌(Corynespora cassiicola)对QoI类杀菌剂的抗性高达100%。2014—2018年采集自阿根廷布宜诺斯艾利斯的82个小麦褐斑病(Pyrenophora tritici-repentis)均为QoIs抗性菌株,以1 mg/L的嘧菌酯、肟菌酯和唑菌胺酯处理,病菌孢子生长不受抑制。而100 mg/L的嘧菌酯、肟菌酯和唑菌胺酯对病原菌也无法完全抑制。
2.3 鞭毛真菌对QoI类杀菌剂抗性发生现状
2001年,整个欧洲采用合理的QoI类杀菌剂施用方案,并对抗药性进行了严密监测,因此,当时欧洲对葡萄霜霉病的控制整体较好,暂无葡萄霜霉病菌(Plasmopara viticola)对QoI类杀菌剂产生抗性的报道。然而,仅1年后在法国和意大利就检测到霜霉病菌对其产生抗性。2005年,王岩等研究结果表明,烯肟菌酯对我国北京、天津、河北、内蒙古、湖北等地采集的52株黄瓜霜霉病菌EC50范围为0.0030~0.0313 mg/L,平均EC50为(0.0101±0.0031)mg/L,具有较高的抗性风险。2021年,Campbell等在美国佐治亚州3个葡萄园采集的39株葡萄霜霉病菌对嘧菌酯和吡唑醚菌酯全部表现出抗性。随着QoI类杀菌剂在生产上的频繁使用和用药量不断上升,抗药性问题呈现加重趋势,延缓和治理QoI类杀菌剂的抗性、延长药剂使用寿命已经成为一个迫切需要解决的科学问题。韩秀英等研究表明,嘧菌酯对19株我国山东、河北、武汉等地采集的黄瓜霜霉病菌的EC50范围为0.00012~0.00934 mg/L,同时建立了黄瓜霜霉病菌对嘧菌酯的敏感基线。赵建江等研究发现番茄叶霉病菌对嘧菌酯存在较高的抗性风险。
3 甲氧基丙烯酸酯类杀菌剂抗性分子机制
3.1 靶标位点氨基酸取代
在大多数情况下,QoI类杀菌剂抗性是由线粒体细胞色素b(cytb)基因的点突变产生。由于细胞色素b由突变率很高的线粒体基因组编码,因此病原菌极易因该基因的突变而导致耐药性发生,在田间极易产生抗药性。
病原菌对QoI类药剂产生抗性的原因主要与cytb基因的127~147位和275~296位间的氨基酸区域有关,主要表现为第143位的甘氨酸变为丙氨酸(G143A),和第129位的苯丙氨酸变为亮氨酸(F129L)。这种氨基酸转换导致病原菌对杀菌剂的敏感性显著降低,并且没有或几乎没有适合度损失,根据病原体和检测系统的不同,抗性系数(RF)为100~1,000。具F129L的菌株大都表现中低抗性,相比之下,G143A分离株基本表现出完全高抗性,且出现的频率更高,例如在欧洲、澳大利亚和美国多个州,从小麦白粉病菌、小麦叶斑病菌、小麦颖枯病菌、大豆灰斑病菌中均鉴定出G143A突变。目前,Neves等建立了一种基于G143A突变的PCR检测方法,可以成功区分QoI敏感和耐药菌株。准确监测这一突变将有助于减缓QoI抗性的传播,并对这一病原菌系统中的杀菌剂抗性管理具有重要意义。
此外,在小麦黄斑叶枯病菌(Pyrenophora tritici-repentis)中还发现了G137R的位点突变,Cosseboom等在葡萄晚季腐烂病菌中发现了包含S108A和A194V的突变,但这2种突变的抗性都较低。Zhou等在荔枝露疫病菌的抗性株中发现G142A、G142S、Y131C、F128S等突变,通过分子对接技术分析表明,由于药剂自身结构的差异及与口袋腔作用力变化强弱的程度不同,F128S、Y131C突变株对于不同的杀菌剂具有RF>500和RF<100不同程度的抗性。
3.2 旁路氧化途径
除了cytb基因中靶标位点氨基酸的取代,其他机制也可能诱导病原真菌对QoIs的耐药性。交替氧化途径过程中的关键酶AOX是一种单亚基(非血红素)含双铁的单分子膜蛋白,广泛存在于植物、真菌和原生线粒体中,其在呼吸过程中充当复合物Ⅲ和Ⅳ的旁路,形成一种不同于细胞色素系统的电子传递系统,在植物病原体线粒体内形成另一种呼吸途径,即线粒体的电子通过绕过细胞色素bc1复合物QoI的抑制位点而转移,这种旁路有效克服了QoI类杀菌剂对复合物Ⅲ的抑制作用。
在亚致死浓度的QoI杀菌剂压力下,交替呼吸可能是植物病原真菌从敏感到全面抗性转变的重要途径。AOX有诱导表现型和组成型2种形式,前者多发生在以细胞色素为介质的呼吸途径被阻断或线粒体蛋白的合成受到抑制的情况下,AOX被诱导表达,后者则是不受呼吸途径的影响,不需要其他因子的诱导即可表达,一般对甲氧基丙烯酸酯类杀菌剂的敏感性比诱导型的敏感性差。在植物体外,AOX活性可以被旁路氧化酶抑制剂水杨肟酸(SHAM)抑制,SHAM是一种相对活性较低的AOX抑制剂,其作为一种抗真菌药物被观察到在体外与QoI类杀菌剂具有协同增效作用。杨维等在含SHAM培养基中测定肟菌酯对香蕉黑斑病菌菌丝生长的抑制,结果表明,水杨肟酸存在时,肟菌酯对菌丝生长的抑制比肟菌酯单独作用效果明显,但在抗药菌株中,未观察到明显区别,说明交替氧化途径在敏感菌株中更为重要。
3.3 外排转运体机制
ABC(ATP-binding cassette)和MFS(major facilitator superfamily)复合体蛋白是最具代表性的保护病原菌免受杀菌剂影响的外排转运体,可转运多种底物,介导各种有毒底物从细胞外流,防止毒素积累。合成杀菌剂通常被认为是对真菌种群的毒素,其功效因此会受到真菌体内药物外排转运体活性的影响。植物病原菌ABC转运蛋白的研究主要集中在参与植物对杀菌剂的多重抗性,如灰霉病菌中的BcATRB和BcATRD以及青霉菌中的PMR1和PMR5。已有大量研究报道了ABC转运体在病原菌多药抗性(multi-drug resistance)中发挥着重要作用。研究者在构巢曲霉中第一次检测到由atrB基因编码的ABC转运体与QoI类杀菌剂抗性存在相关性,发现该基因过表达可以保护病原菌免受几乎所有供试杀菌剂的抑制作用。MFS转运蛋白在病原菌的多药抗性中也发挥着重要作用,小麦叶枯病菌的多药抗性相关研究指出,Mgmfs1基因的启动子序列可发生519bp的插入突变,致使该基因编码的转运蛋白过量表达,从而导致其产生多药抗性。有报道称通过插入编码基因的启动子可诱导转运蛋白MFS1的过表达,由此引起病原菌产生多重耐药性。
4 植物病原菌对QoI类杀菌剂抗药性治理
甲氧基丙烯酸酯类杀菌剂在全世界受到广泛的应用,已超越三唑类成为全球第一大杀菌剂类型,可防治不同作物的多种病害。抗性数据分析表明,该类杀菌剂存在着较高的抗性风险。不合理使用药剂会使抗药性突变菌株在较短时间内形成抗药性群体,导致杀菌剂失效,从而造成严重损失,如在西班牙的穆尔西亚瓜类种植区,仅在2008年瓜类白粉病菌(Podosphaera fusca)中多达74%的菌株对QoI类杀菌剂已产生了抗性。因此,治理抗药性必须合理规范药剂的使用。
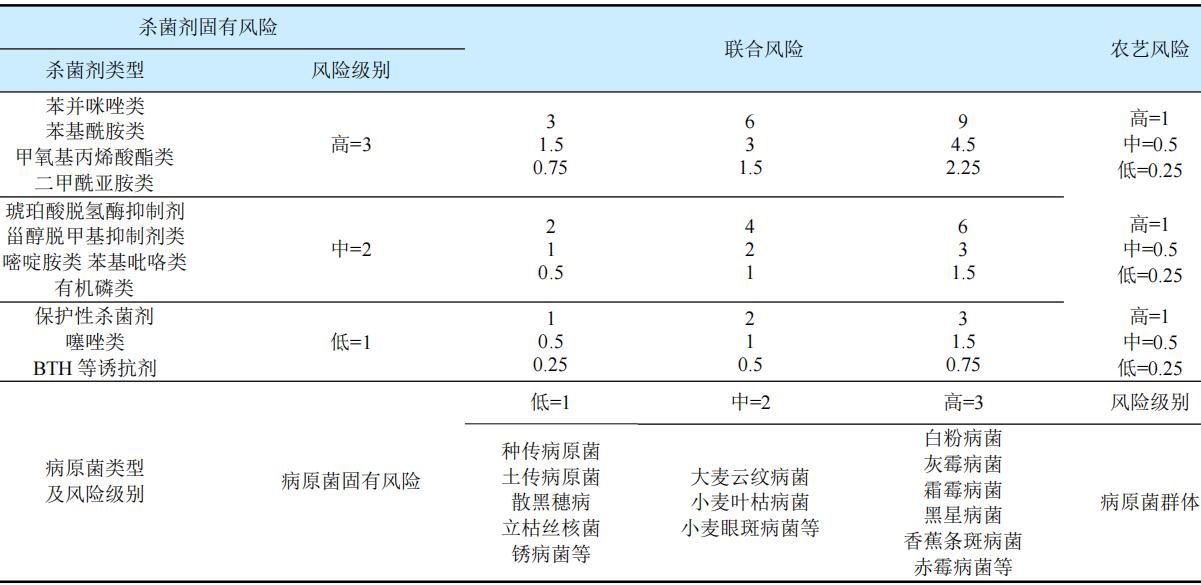
抗性风险管理需要多方合作,包括但不限于3个方面:降低杀菌剂本身固有风险、规范杀菌剂施用者操作以及农业相关部门的积极管理。杀菌剂抗性行动委员会(FRAC)将病原菌固有风险和杀菌剂固有风险划分为高、中、低3个等级(1=低,2=中,3=高),将农艺风险等级同样划分为3个等级(0.25=低,0.5=中,1=高),结合病原菌固有风险、药剂固有风险和农艺风险等级,以三者的乘积作为相对应病原菌对杀菌剂的抗性风险等级(联合风险)。理论抗性风险划分标准:0~1=低等风险,1~3=中低风险,3~6=中风险,6=中高风险,9=高风险(表1)。因此,首先应尽量减少利用甲氧基丙烯酸酯等高风险类杀菌剂防治同样为高风险的病原菌,如白粉病菌、灰霉病菌、霜霉病菌等,避免产生较高的联合风险。其次,杀菌剂的轮换和混配使用是延缓抗性的有效手段之一,避免长期单一使用某一产品或某一类药剂,必须采用科学的混配用药和轮换用药防治措施,即与不同作用机理或不存在交互抗性的杀菌剂混用或轮换用药,例如图1中C3(Group 11)的杀菌剂之间存在交互抗性,而C3(Group 11A)中的杀菌剂与存在G143A突变的C3(Group 11)中的杀菌剂无交互抗性,因而可使用C3(Group 11A)中的杀菌剂与之进行混配使用。
表1 杀菌剂固有抗性风险和病原菌固有抗药性风险等级
在此基础上,可进一步研制开发具有增效作用或具有协同作用的杀菌剂混剂,根据抗药病原群体形成的主要影响因素,针对性地设计抗药性治理策略。综合考虑所有与抗药性发生相关的影响因子,抗药性的治理也应当从施药者因素出发。进一步建立完善一系列正确的施药操作规范,加大全方位宣传教育,落实到每一位相关从业者,使其明确科学合理用药的重要性,不得私自加大施药量。企业与产品销售所在地方县级农业管理部门应积极合作,组织开展对农药经营者和农药施用者进行抗药性治理、科学用药及实际操作性知识培训。例如,施药者应在病害早期施用杀菌剂,避免在病害严重发生时期喷施高剂量的药剂,一则达不到有效的防治,二则在高浓度药剂的定向选择下,加速了田间抗性群体的产生和发展。同时,施药者还需掌握一定的农业防治措施,如利用非同一寄主作物进行轮作或休耕,改变播期,降低田间湿度,注意田园清洁,及时移除、深埋或焚烧修剪或摘除病花、病果、病叶等作物组织等。同时,选用物理防治、生物防治等其他有利于减轻病害发生和危害的非化学防治措施,例如选用抗病品种,增强植物对病害的抗病性。该措施旨在精准有效防控病害,减少农药用量,防止病害突发性流行危害,促进农业增产与农民增收。此外,为延缓该类药剂抗药性突变体的产生速度,对于已有报道显示对甲氧基丙烯酸酯类杀菌剂产生抗药性的作物病害,农业农村部农药登记管理部门应适当控制防治该病害具有交互抗性的药剂登记数量,并组织登记同类产品的企业开展抗药性监测。在登记药剂的产品标签上标注抗性风险等级,并标明相应的抗性风险管理措施、生长季该产品的使用次数、不能与其他哪类药剂混用或轮用等信息。在杀菌剂推广应用之前,药企必须具有抗药性风险评估报告,预测病原菌产生抗药性的潜在风险,并在产品标签上标注抗性风险级别。这有助于在田间出现实际抗药性导致药剂防效下降之前,指导生产中及早采用预防性的抗药性治理。
与此同时,日本杀菌剂抗药性行动委员会(Japan FRAC)建议,在梨上每年最多使用2次QoI类杀菌剂,每个季节使用吡菌苯威的次数也需要少于3次;欧洲农业政策指出,从2014年起,所有欧盟成员国都应实施虫害综合治理(IPM),以减少农药的影响和使用。此外,开展病原菌对QoI类杀菌剂的抗性监测和鉴定工作在病害防治中十分重要。大量文献强调细胞色素b靶标位点氨基酸取代在QoI类杀菌剂抗性中的作用,但随着近年来对QoI类抗性机制的认识不断深入,越来越多的研究将QoI类抗药现象归因于不同于靶标位点氨基酸取代的机制,例如交替呼吸、外排转运体和其他等未知机制。因此检测病原菌群体抗药性不能仅仅依据cytb靶标位点氨基酸取代,还需建立杀菌剂和防治对象的敏感性基线和抗性监测方法,根据监测结果,明确病原菌对杀菌剂抗药性的产生和发展情况,在实践中进一步调整、补充和完善抗性治理策略,通过改变杀菌剂的应用方案,从而有助于防止对QoI类耐药的病原体群体的进一步发展。
(1)本网旨在传播信息,促进交流,多方面了解农药发展动态,但不构成任何投资建议。
(2)所有文章仅代表作者观点,不代表本网立场。
(3)“信息来源:江苏省农药协会 农药资讯网”为原创文章,转载时请注明来源和作者。
(4)本网转载文章及图片的版权属于原作者,若有侵权,请联系删除。