




棉蚜Aphis gossypii Glover,又称瓜蚜,属半翅目蚜科,是一种世界性分布的重要害虫。棉蚜能够为害棉花、木槿、黄瓜、甜瓜、茄子、辣椒和小麦等300多种植物。作为一种重要的刺吸式口器害虫,棉蚜可通过直接取食植物汁液和传播植物病毒造成危害。长期以来,防治棉蚜主要依赖于化学杀虫剂,但棉蚜具有种群数量大、繁殖速度快、适应性强等特点,能快速适应防治药剂并产生抗药性,严重影响化学防治的效果,对我国棉花高质量安全生产造成严重的经济损失。新烟碱类杀虫剂是近年来棉蚜田间防治的主要药剂,但棉蚜对该类药剂产生较高水平的抗性,已成为棉蚜有效防控中面临的最大挑战。本文从棉蚜对新烟碱类杀虫剂的抗性现状、抗性机制以及抗性治理策略等方面进行综述,以期为棉蚜抗药性科学治理提供参考。
1 新烟碱类杀虫剂
新烟碱类杀虫剂是全球最重要的一类化学杀虫剂,对多种害虫具有较高的杀虫活性,是防治蚜虫、飞虱、叶蝉等刺吸式口器害虫最有效的杀虫剂种类之一。尽管早在1690年人们就开始用烟草浸出液(主要成分为烟碱)防治农业害虫,但新烟碱类杀虫剂的正式研制始于1970年壳牌公司以烟碱结构为基础的先导化合物SD-031588的开发。1985年,德国拜耳公司和日本农药株式会社合作开发出了第一个新烟碱类杀虫剂——吡虫啉,并于1991年由拜耳公司进行全球推广。随后,国际上的几家大型农药公司相继研发出烯啶虫胺、啶虫脒和噻虫嗪等多种新烟碱类杀虫剂(图1)。
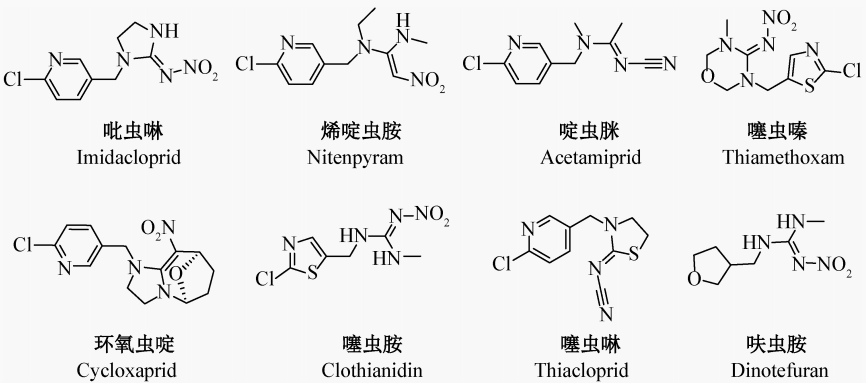
图1 新烟碱类杀虫剂
新烟碱类杀虫剂作用于昆虫烟碱型乙酰胆碱受体(nicotinic acetylcholine receptor,nAChR),是nAChR的选择性激动剂,该类杀虫剂能够通过与昆虫nAChR的结合阻断昆虫中枢神经系统信号的正常传导,导致害虫死亡。新烟碱类杀虫剂具有高效、广谱、内吸性好的特点,兼具触杀和胃毒作用,对哺乳动物低毒,可有效防治半翅目、鞘翅目、双翅目等多种害虫。自吡虫啉上市以来,新烟碱类杀虫剂因其良好的杀虫活性,在世界范围内已成功取代了有机磷类、氨基甲酸酯类等传统药剂在作物保护中的应用。新烟碱类杀虫剂在全球市场中占有重要地位,在2014年该类杀虫剂的市场占有率在25%以上,近年来其在全球杀虫剂市场中的占有率虽有下降,但仍是全球第一大杀虫剂类型。
2 棉蚜对新烟碱类杀虫剂的抗性现状
2.1 国外抗药性现状
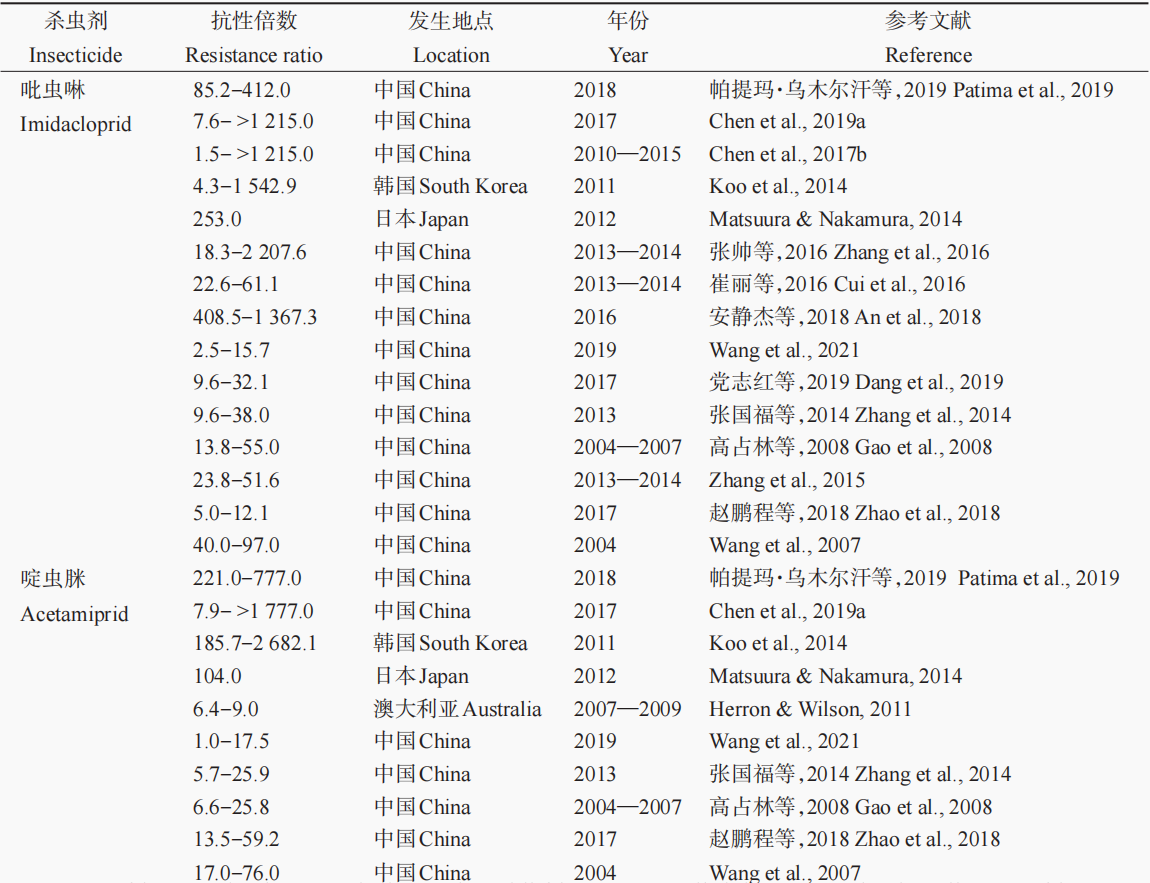
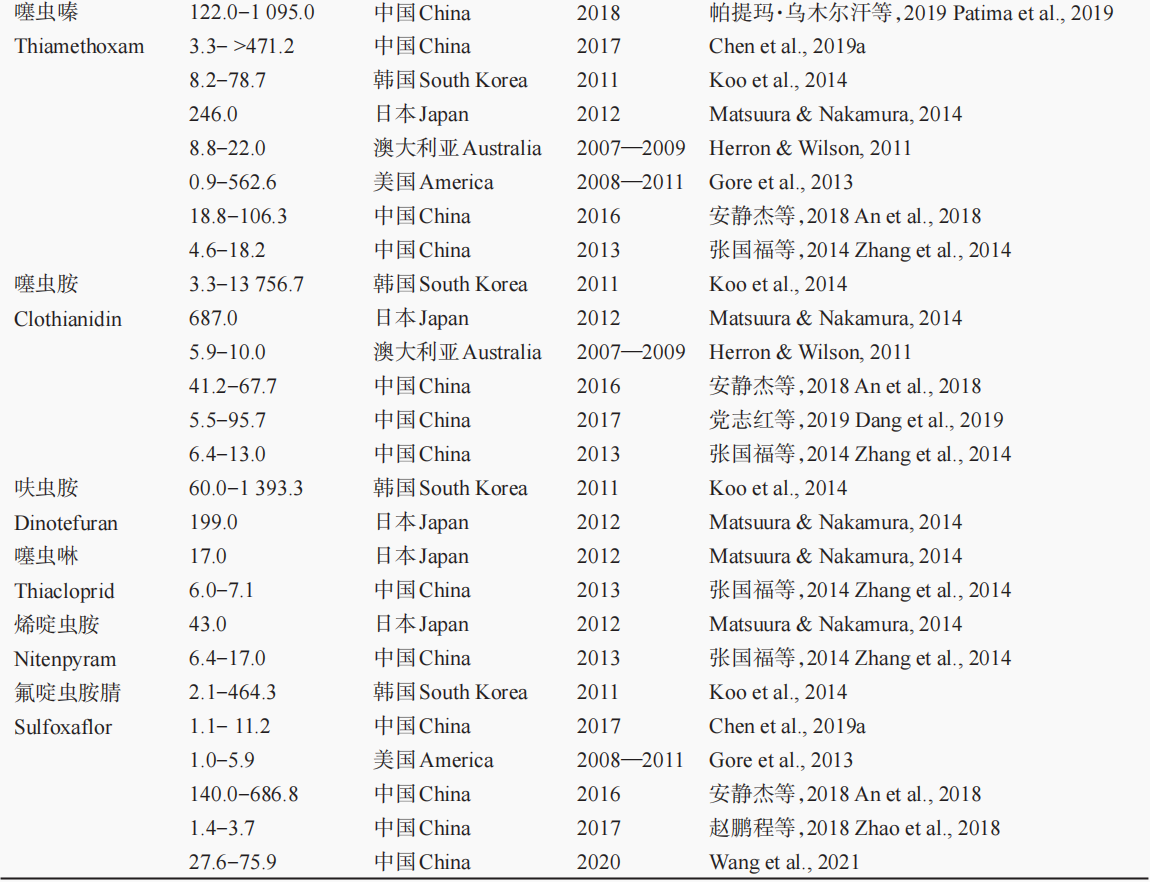
随着新烟碱类杀虫剂使用量的增加,早在2005年害虫对新烟碱类杀虫剂的抗性问题就引起了人们的关注。棉蚜作为一种抗性进化很快的农业害虫,已对多种新烟碱类杀虫剂产生了抗性。尽管在20世纪90年代新烟碱类杀虫剂才开始市场化推广,但在2000年左右就有文献报道棉蚜对新烟碱类杀虫剂产生了抗性(表1)。自2001年首次发现意大利和葡萄牙棉蚜种群对吡虫啉敏感性降低以来,棉蚜对新烟碱类杀虫剂的抗性问题日益严重。例如,Herron & Wilson通过诊断计量法监测发现,2006—2008年澳大利亚棉蚜田间种群对新烟碱类杀虫剂的抗性逐年加重,至2008年已对啶虫脒、噻虫胺和噻虫嗪分别产生了6.4倍、10倍和22倍的抗性,且这种抗性趋势还在不断发展,到2009—2010年所监测的澳大利亚棉蚜种群中抗性个体的比例已经高达96%。在美国,2008—2011年的抗药性监测结果表明,棉蚜对噻虫嗪已经产生了高水平抗性,其抗性倍数最高已达562.6倍。2012年,在日本宫崎市也发现采自黄瓜和甜椒上的棉蚜种群对吡虫啉、呋虫胺、噻虫胺、噻虫嗪、烯啶虫胺、啶虫脒和噻虫啉这7种新烟碱类杀虫剂均产生了中等至高等水平的抗性,其中对噻虫胺的抗性水平最高为687倍,对噻虫啉抗性水平最低也达到17倍,而对其余5种新烟碱类杀虫剂的抗性倍数在43~253倍之间。同样,在韩国,2012年的抗性监测结果也表明,采自辣椒和黄瓜上的棉蚜对6种新烟碱类杀虫剂均产生了较高水平的抗性,其中对啶虫脒、噻虫胺、吡虫啉和噻虫啉的抗性倍数最高分别达2,682倍、13,756倍、1,542倍和1,393倍。以上研究结果表明,棉蚜对新烟碱类杀虫剂的抗性十分严重,已成为世界性难题。
表1 棉蚜抗药性现状
2.2 国内抗药性现状
国内棉蚜种群对新烟碱类杀虫剂的抗性报道稍晚于国外(表1)。2004年,Wang等监测发现采自山东省德州市、菏泽市、聊城市、德州市、泰安市等地的棉蚜田间种群对吡虫啉和啶虫脒分别产生了40~97倍和17~76倍的抗性。2004—2007年,河北省廊坊市、保定市、沧州市等地的棉蚜种群对吡虫啉的抗性均达到了中等至高等水平,最高抗性倍数达55倍;廊坊市和沧州市棉蚜种群对啶虫脒也产生了低等至中等水平抗性,抗性倍数最高达25.8倍。2013年,张国福等对采自山东省聊城市、东营市和泰安市的棉蚜种群对6种新烟碱类杀虫剂的抗性水平进行监测,发现山东省棉蚜种群对吡虫啉、啶虫脒、烯啶虫胺、噻虫嗪和噻虫胺均产生了中等水平抗性,对噻虫啉产生低等水平抗性。2013—2014年,邱县、滨州市、西华县的棉蚜种群对吡虫啉的抗性倍数均大于900倍,达到高水平抗性。张学涛等于2010年的监测结果表明,新疆维吾尔族自治区(简称新疆)石河子市的棉蚜田间种群对啶虫脒产生了高水平抗性,抗性倍数最高达270.0倍。河北省廊坊市、新疆阿克苏、山东省德州市和新疆奎屯市棉蚜种群对吡虫啉分别产生了22.6倍、26.3倍、53.5倍和61.1倍的抗性。2018年,帕提玛·乌木尔汗等研究发现,采自新疆棉区的棉蚜种群对新烟碱类杀虫剂产生了中、高水平的抗性,其中对吡虫啉、啶虫脒和噻虫嗪的抗性分别为85.2~412倍、221~777倍和122~1,095倍。同年,安静杰等监测结果表明,河北省棉蚜种群对吡虫啉、噻虫嗪、噻虫胺均产生了中等至高等水平抗性,其中对吡虫啉的抗性水平高达1,367.3倍。2019年,部分采自山东省黄瓜上的瓜蚜(棉蚜)种群对吡虫啉和啶虫脒产生了中等水平抗性,抗性倍数最高分别达15.7倍和17.5倍。以上结果表明,我国棉蚜田间种群对新烟碱类杀虫剂的抗性水平普遍较高,需继续加强抗性治理,以延缓抗性发展,提高新烟碱类杀虫剂对棉蚜的防治效果。
2.3 交互抗性
新烟碱类杀虫剂因其与其他种类杀虫剂无交互抗性而被广泛使用,但越来越多的研究发现新烟碱类杀虫剂之间存在较高的交互抗性风险,值得大家注意。例如,Chen等交互抗性测定结果表明,抗吡虫啉棉蚜品系(抗性倍数>1,215倍)对啶虫脒和氟啶虫胺腈具有高达1,776倍和260倍的交互抗性。Wei等发现抗噻虫嗪棉蚜品系对联苯菊酯(11.71倍)、氟氯氰菊酯(17.90倍)、氰戊菊酯(6.85倍)、噻虫胺(6.56倍)、杀扑磷(5.34倍)和高效氯氰菊酯(4.53倍)均存在明显的交互抗性。Zhang等测定结果也表明,抗噻虫嗪瓜蚜(棉蚜)品系(抗性倍数>2,325.6倍)对啶虫脒、呋虫胺、烯啶虫胺和噻虫胺分别具有76.6倍、142.3倍、299.9倍和311.7倍的交互抗性。此外,抗呋虫胺瓜蚜(棉蚜)品系(抗性倍数为74.7倍)对噻虫嗪具有15.3倍的交互抗性。
3 棉蚜对新烟碱类杀虫剂的抗性机制
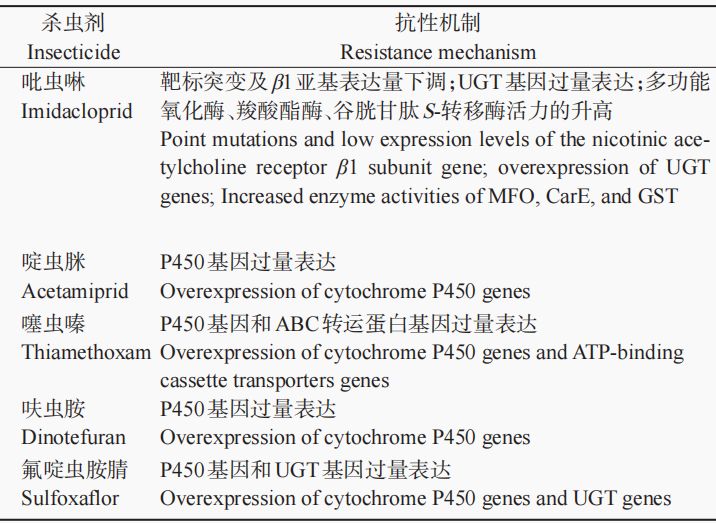
棉蚜对新烟碱类杀虫剂产生抗性的机制主要涉及解毒代谢能力的增强和靶标敏感性的下降(表2),下面将主要从细胞色素P450、谷胱甘肽S-转移酶等介导的代谢抗性和靶标抗性两方面进行论述。
3.1 代谢抗性
3.1.1 细胞色素P450介导的抗药性
细胞色素P450是生物体内最重要的解毒代谢酶,在害虫抗药性产生过程中具有非常重要的作用。作为多功能氧化酶系的主要组成部分,细胞色素P450对杀虫剂的代谢能力直接影响昆虫对杀虫剂的耐受性。P450介导的杀虫剂代谢解毒作用是许多害虫对杀虫剂产生抗性的重要机制。目前已知棉蚜对新烟碱类杀虫剂的抗性大多与P450介导的解毒代谢增强有关。例如,石绪根通过增效剂试验和酶活性测定试验,发现多功能氧化酶在棉蚜对吡虫啉的抗性中起着重要作用。郭天凤等同样发现多功能氧化酶与棉蚜对吡虫啉和啶虫脒的抗性密切相关。Wu等通过对P450基因表达量的分析发现,CYP3家族P450基因的上调表达可能是棉蚜对噻虫嗪产生抗性的主要原因。同样,CYP6CY14、CYP6DC1和CYP6CZ1等P450基因的过表达是棉蚜对啶虫脒产生代谢抗性的重要机制,CYP6CY14、CYP6CY22和CYP6UN1的过表达在棉蚜对呋虫胺产生抗性过程中起重要作用。此外,Hirata等通过S2细胞真核表达在新烟碱类杀虫剂抗性品系中显著上调表达的CYP6CY22和CYP6CY13基因,并进行体外代谢试验,发现棉蚜P450能够快速代谢吡虫啉、啶虫脒、噻虫啉等新烟碱类杀虫剂,证实了P450基因过表达是介导棉蚜对新烟碱类杀虫剂产生代谢抗性的重要机制。
3.1.2 羧酸酯酶介导的抗药性
羧酸酯酶作为另外一种十分重要的解毒酶,能够催化水解含羧基酯键、酰胺键和硫酯键的化合物。许多用于棉蚜防治的化学杀虫剂,如有机磷类、氨基甲酸酯类、拟除虫菊酯类等杀虫剂,其分子结构中都含有酯键,因此,羧酸酯酶代谢能力的增强也是棉蚜抗药性产生的重要机制之一。已有研究表明,羧酸酯酶活性(以αNA为底物)在棉蚜吡虫啉抗性品系中均显著高于敏感品系,推测其与棉蚜对吡虫啉的抗性有关。杨焕青等研究发现,对吡虫啉产生24.38倍抗性的棉蚜品系的羧酸酯酶比活力是敏感品系的2.73倍,史晓斌等也发现吡虫啉抗性棉蚜品系的羧酸酯酶比活力是敏感品系的2.87倍,说明棉蚜对吡虫啉的抗性产生与羧酸酯酶解毒能力的升高有关。崔丽等对田间棉蚜抗性品系的酶活性测定结果表明,对吡虫啉产生中等水平抗性的棉蚜田间种群(阿克苏种群和奎屯种群)羧酸酯酶的比活力分别是敏感种群的1.6倍和1.7倍,说明羧酸酯酶活力的升高与棉蚜对吡虫啉的抗性形成有关。值得注意的是,新烟碱类药剂的化学结构中不含酯键,那么羧酸酯酶如果作为吡虫啉的抗性机制的话仅有一种可能性,即作为结合蛋白起作用。但是上述研究报道仅是一个相关性,并没证明作为结合蛋白的作用。因此,羧酸酯酶介导棉蚜对新烟碱类杀虫剂抗性的分子机制并不明确。
3.1.3 谷胱甘肽S-转移酶介导的抗药性
谷胱甘肽S-转移酶催化谷胱甘肽结合反应最开始的步骤,是该反应的关键酶,催化亲核性(还原性)的谷胱甘肽与不同种类的亲电子外源化合物发生反应,使还原性谷胱甘肽与有害亲电子基团结合并排出体外,起到解毒代谢的作用。以谷胱甘肽为介质的代谢,主要有2种:第1种是以还原型谷胱甘肽作为催化剂的代谢反应,第2种是还原型谷胱甘肽与底物直接结合的代谢反应。有研究表明谷胱甘肽S-转移酶与棉蚜对新烟碱类杀虫剂抗性相关,例如,潘文亮等通过酶活性测定试验证实谷胱甘肽S-转移酶活力增强在棉蚜对吡虫啉的抗性中起重要作用。杨焕青等(2009)研究也发现,抗吡虫啉棉蚜品系谷胱甘肽S-转移酶的比活力是敏感品系的1.57倍,说明谷胱甘肽S-转移酶比活力的提高是引起棉蚜对吡虫啉产生抗性的重要原因之一。崔丽等认为,我国新疆阿克苏地区和奎屯市的棉蚜田间种群对吡虫啉的抗性与谷胱甘肽S-转移酶活性的增强相关。
3.1.4 UDP-葡糖基转移酶介导的抗药性
UDP-葡糖基转移酶(UDP-glycosyltransferase,UGT)是一种重要的Ⅱ相代谢酶,能够催化多种内源和外源物质与葡萄糖醛酸结合,在昆虫内源和外源化合物的解毒代谢中具有重要作用。近年来,有研究表明UGT在害虫中起非常重要的解毒代谢作用,参与害虫对防治药剂的抗性形成。例如,Li等研究发现UGT在小菜蛾Plutella xylostella(L.)对氯虫苯甲酰胺抗性形成中起着重要作用。在棉蚜中,UGT也被发现在棉蚜对新烟碱类杀虫剂的抗性产生过程中起着重要的解毒代谢作用。例如,在棉蚜对噻虫嗪抗性品系中有13个UGT基因显著上调表达,且RNAi沉默UGT348A2、UGT344B4和UGT344J2这3个UGT基因表达均能显著提高抗性棉蚜对噻虫嗪的敏感性,说明UGT基因的过量表达与棉蚜对噻虫嗪的抗性相关。Chen等研究发现,在棉蚜吡虫啉抗性品系中UGT343A4、UGT344B4、UGT344C7等多个UGT基因显著上调表达,且在蛋白水平上UGT344A16、UGT344B4、UGT344C7和UGT344N4的表达水平也显著高于敏感品系,进一步RNAi沉默UGT344B4和UGT344C7的表达能够显著提高吡虫啉抗性品系棉蚜对吡虫啉的敏感性,证实UGT344B4和UGT344C7的过量表达与棉蚜对吡虫啉的抗性有关。Ma等在棉蚜对氟啶虫胺腈抗性机制研究中也发现,UGT344B4、UGT344C5、UGT344A11等7个UGT基因在棉蚜氟啶虫胺腈抗性品系中显著上调表达,而RNAi沉默4个UGT344亚家族基因(UGT344B4、UGT344C5、UGT344A11和UGT344L2)均能够显著提高棉蚜对氟啶虫胺腈的敏感性,说明多个UGT基因的过表达也是棉蚜对氟啶虫胺腈产生代谢抗性的重要原因之一。
3.1.5 ABC转运蛋白介导的抗药性
Pan等研究发现,与敏感品系相比,棉蚜对噻虫嗪抗性品系中多个ABC转运蛋白基因显著上调表达,且抑制过表达的ABC转运蛋白基因ABCA2、ABCD1、ABCD2、ABCE1和ABCG15能够显著提高抗性棉蚜对噻虫嗪的敏感性,说明棉蚜对噻虫嗪的抗性与ABC转运蛋白有关。类似地,Wang等通过比较室内敏感品系和氟啶虫胺腈抗性品系ABC转运蛋白基因的表达差异,发现AB‐CA1、ABCA2、ABCB1等10个ABC转运蛋白基因在抗性品系中显著上调表达,且RNA干扰ABCA1和ABCD1两个基因显著提高了抗性棉蚜对氟啶虫胺腈的敏感性,说明ABC转运蛋白基因的上调表达也参与了棉蚜对氟啶虫胺腈的抗性形成。
3.2 靶标抗性
分子靶标抗性变异是害虫产生抗药性的重要机制。新烟碱类杀虫剂的作用靶标是烟碱型乙酰胆碱受体,因此,nAChR靶标位点突变是棉蚜对新烟碱类杀虫剂产生抗性的重要机制。Koo等研究发现棉蚜烟碱型乙酰胆碱受体β1亚基环D区的R81T突变是其对吡虫啉产生抗性的重要机制,且体外试验证实R81T突变介导的靶标敏感性下降与棉蚜对新烟碱类杀虫剂的抗性直接相关。Kim等在棉蚜吡虫啉抗性品系中检测到L80S和R81T突变,也证实其与吡虫啉抗性相关。Zhang等采用实时荧光定量特异性等位基因PCR(real-time PCR amplififi cation of specifific-allele,RT-PASA)技术检测发现,采自廊坊市和德州市的田间吡虫啉抗性棉蚜种群中存在R81T突变个体,而崔丽等通过对我国田间棉蚜种群的突变检测也发现,nAChR β1亚基的R81T突变与棉蚜对吡虫啉的抗性相关。Chen等在室内筛选的吡虫啉高抗品系(抗性倍数>1,215倍)的β1亚基上同时检测到了R81T、V62I和K264E三个位点突变,其中V62I和K264E突变为首次发现,V62I位于LoopD前翼序列,而K264E位于跨膜区域TM1和TM2之间,进一步对田间种群的检测结果也证实这3个位点突变均与棉蚜对吡虫啉的抗性相关,说明nAChR靶标突变是棉蚜对吡虫啉产生抗性的关键原因。此外,Munkhbayar等和Wang等通过对采自我国新疆和山东省的棉蚜田间种群的突变检测也发现nAChRβ1亚基上R81T和K264E突变与棉蚜对新烟碱类杀虫剂的抗性有关。
此外,nAChR受体基因低水平表达可能也在棉蚜对新烟碱类杀虫剂的抗性形成中发挥着重要作用。例如,比较吡虫啉抗性和敏感品系nAChRβ1亚基的表达量,发现其表达量在棉蚜吡虫啉抗性品系中显著下调,说明其低水平表达与棉蚜对吡虫啉的抗性形成有关。同样,在棉蚜噻虫嗪抗性品系中,在nAChR各亚基中均未检测到靶标位点突变,但其α1、α4-1、α4-2、α5及α7亚基在转录水平上分别下调表达3.32倍、1.60倍、2.05倍、5.41倍和1.48倍,说明棉蚜α1、α4-1、α4-2、α5及α7亚基的下调表达可能在棉蚜对噻虫嗪的抗性形成中起着一定作用。
表2 棉蚜对新烟碱类杀虫剂的抗性机制
4 棉蚜抗性治理策略
Georghiou & Saito从化学药剂使用的角度提出了害虫抗药性治理的3种策略,即适度治理、饱和治理和多向进攻治理。适度治理主要通过降低杀虫药剂的选择压力,延缓抗性发展。具体措施包括采用低剂量用药,保留一定比例的敏感基因型,减少施药次数,选用残效期短的化合物,避免使用缓释剂,主要针对成虫施药,尽量局部而不是大面积施药,人为设置一定的庇护区,提高施药害虫种群阈值等。和适度治理相反,饱和治理的目的是尽可能的提高选择压,消除抗性杂合子中的抗性等位基因,延缓抗性发展。其主要措施是通过高剂量用药,杀死大多数杂合子,使抗性功能隐性化。多向进攻治理主要是根据毒物对生物的多位点作用,使靶标不易产生抗性,一般通过杀虫药剂的轮用、混用以及分区施药来延缓抗性发展。针对棉蚜抗性严重的特点,建议采取以下几种措施,尽量延缓棉蚜的抗性发展。
4.1 加强田间种群抗药性动态监测
抗药性监测是制定害虫抗药性治理策略和精准选药的关键,要综合利用各种抗药性检测技术,系统测定棉蚜对新烟碱类杀虫剂的抗性发生频率和抗性水平的时空变化,了解其抗性发生和发展的规律。为尽量延长新杀虫剂的使用寿命,应该从其开始应用之时就制定抗性监测计划,并持续进行监测。同时,利用现代分子生物学技术,调查棉蚜对新烟碱类杀虫剂抗性形成的相关基因资源,了解抗性相关基因的种类、频率及其分布。根据抗性监测结果,制定抗性治理策略,指导田间科学用药,延缓棉蚜对新烟碱类杀虫剂的抗性发展,延长杀虫剂的使用寿命。
4.2 合理选择混用和轮用杀虫剂组合
抗性治理中的药剂混用是利用一组具有独立作用机制的化合物,形成多位点作用,其中任何一种化合物对昆虫的选择压力都低于抗性发展的要求,因而不会引发抗性的产生。杀虫剂混用防治害虫时,一种组分不能杀死的个体可能被另一组分杀死,因为对所有杀虫剂都具有抗性的个体几乎不存在。杀虫剂混用作为克服或延缓抗性发展的措施需注意以下几点:① 混用的各组分的作用机制应彼此不同,如果作用机制相同,则害虫很可能对混剂中的各组分产生交互抗性,反而促进了抗性发展;② 害虫对混用的每一组分的抗性机制应该不同,即害虫对混用各组分的抗性机制不同;③ 混用的各组分的残效期应基本相等,如果混用的组分之一残效期明显长于其他组分,将会造成选择压力不平衡,失去混用的意义。
杀虫剂轮用的主要依据是害虫抗药性是在杀虫剂存在条件下“瞬间进化”的结果,但抗性个体在对杀虫剂产生抗性的同时往往会产生适合度代价,如发育期延长、生殖力下降、个体变小、体重减轻等;当产生抗性的“瞬间进化”条件(杀虫剂的选择压力)消失时,抗性个体因为生物学上的劣势而逐渐被淘汰,在种群中的频率下降,最终抗性种群对药剂的敏感度会恢复。进行杀虫剂轮用时要注意:最好选择具有负交互抗性的药剂,以形成反选择作用,有效延缓或阻止害虫抗药性的发生;选用作用机制不同的药剂进行轮用,避免形成交互抗性;设置合理的轮用间隔期,这是杀虫剂轮用能否成功的关键,轮用间隔期一定要足以使昆虫种群对上一次所使用药剂的抗性得到恢复;杀虫剂的轮用同样也应该在药剂开始应用的早期就实施。例如,在棉蚜田间防治过程中可以轮换使用对棉蚜防控效果较好,且与新烟碱类杀虫剂无交互抗性的新型杀虫剂双丙环虫酯,以延缓棉蚜对新烟碱类杀虫剂的抗性发展。
4.3 合理使用增效剂
增效剂是一类本身对害虫没有生物活性或活性较低的化合物,但其与杀虫剂联合使用后会使杀虫剂的毒力显著增强。增效剂可以通过抑制相关解毒代谢酶的活性,消除因代谢抗性机制产生的抗性,从而达到提高药效,减少用药量,延缓抗性发展的目的。代谢抗性是棉蚜对新烟碱类杀虫剂产生抗性的重要原因,因此合理使用增效剂能够在一定程度上延缓棉蚜对新烟碱类杀虫剂的抗性发展。例如,郭天凤等发现,增效醚(piperonyl butoxide,PBO)和三苯基磷酸酯(triphenyl phosphate,TPP)能够显著提高棉蚜对吡虫啉和啶虫脒的敏感性;Chen等研究表明,PBO和TPP两种增效剂能够显著提高棉蚜对呋虫胺的毒力。因此,尽管PBO和TPP可能不能直接作为增效剂使用,但是这些研究结果表明通过合理使用增效剂可以达到提高新烟碱类杀虫剂对棉蚜的防治效果,减少用药次数,降低选择压力的目的。
4.4 充分利用各种防治措施实现综合防治
棉蚜具有繁殖速度快、种群数量大、适应性强的特点,能快速适应防治药剂并产生抗性,在田间棉蚜对新烟碱类杀虫剂的抗性发展十分迅速。因此,要从根本上解决棉蚜对新烟碱类杀虫剂的抗性问题,需要大力开发和利用农业防治、生物防治等环境友好型控制策略,根据综合防治的理念对其进行综合防治。尽量降低棉蚜种群数量,减少棉蚜暴发成灾的可能,降低对化学农药的依赖性。
5 展望
目前绝大多数研究仅能证明P450、UGT等代谢酶解毒代谢增强与棉蚜对新烟碱类杀虫剂抗性的相关性,直接的代谢试验证据还比较少,仅个别基因的表达产物完成了代谢试验,因此还需加强对其直接代谢作用的研究。此外,目前虽有P450等抗性相关基因过表达分子调控机制的报道,但棉蚜对新烟碱类杀虫剂抗性形成的相关基因过表达分子调控机制还较少,仍需加强相关研究。
尽管近年来有双丙环虫酯、氟啶虫胺腈等新型杀虫剂应用于棉蚜的田间防治,但新烟碱类杀虫剂仍是我国目前棉蚜防治中应用最多的一类杀虫剂。因此,棉蚜对吡虫啉、啶虫脒等新烟碱类杀虫剂的抗性问题仍不容忽视,需要继续对其抗药性机理进行深入研究。在棉蚜田间防治和抗药性治理过程中,要持续监测棉蚜田间种群的抗药性现状,并根据抗药性监测结果,制定合理的抗药性治理策略,科学指导田间用药,提高田间防治效果。同时,要根据棉蚜对新烟碱类杀虫剂的抗性机制,在充分利用现有防治药剂的基础上,结合农业防治、物理防治和生物防治等综合防治方法,尽量降低新烟碱类杀虫剂对棉蚜的选择压,以延缓抗性发展,达到对棉蚜进行持续防控的目的。
(1)本网旨在传播信息,促进交流,多方面了解农药发展动态,但不构成任何投资建议。
(2)所有文章仅代表作者观点,不代表本网立场。
(3)“信息来源:江苏省农药协会 农药资讯网”为原创文章,转载时请注明来源和作者。
(4)本网转载文章及图片的版权属于原作者,若有侵权,请联系删除。