




我国每年农田杂草危害发生超过9.3亿公顷,由此造成的经济损失达2,200亿元,除草剂的使用挽回了巨大的损失。然而,除草剂抗性问题日益严重,已成为威胁全球粮食安全的关键问题之一。截止2022年11月,全世界已报道515种除草剂抗性杂草生物型,其中267种分布在72个国家,涉及97种作物。我国是世界上杂草抗性危害最为严重的5个国家之一,其中抗性杂草最多的是小麦田和水稻田。按不同类型划分的抗除草剂杂草种类见图1。图1左为按作用模式划分的抗除草剂杂草种类,从图中可以看出,不同靶标杂草抗性数量在逐年增加,其中作用模式为乙酰羟酸合酶的杂草抗性数量最多;由图1右可以看出,莠去津是物种产生抗性最多的一类除草剂,其次是草甘膦。这些抗性杂草危害严重,防治困难,而除草剂新分子靶标的发现有利于缓解抗性杂草这一严重问题。为此,本文在苏少泉和谭效松等总结除草剂分子靶标类型和功能的基础上,回顾了近30年来发现的除草剂分子靶标,并重点介绍分子靶标的生理功能、除草剂作用机制、除草剂作用范围以及靶标分子结构的研究进展;同时简单介绍当前新靶标发现的困境、靶标发现的新技术以及对未来除草剂靶标发现的展望。
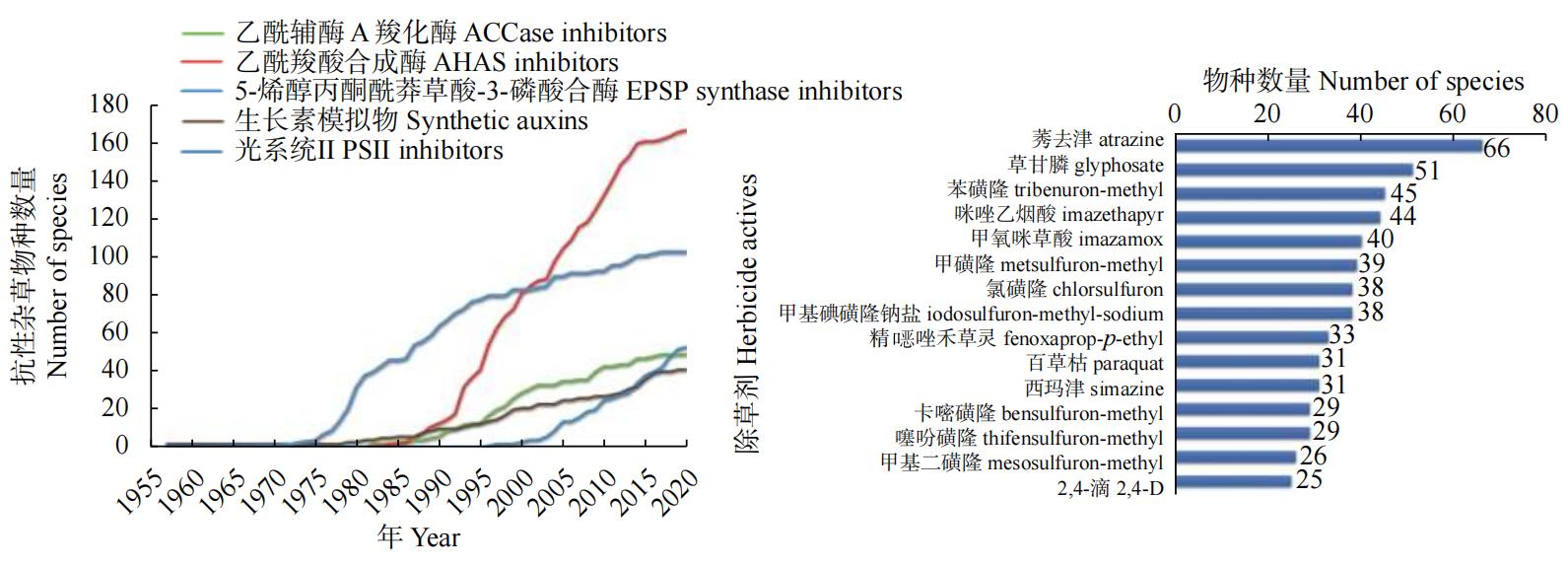
图1 代表性除草剂及代表性作用靶点的抗性统计
1 近30年的除草剂分子靶标汇总
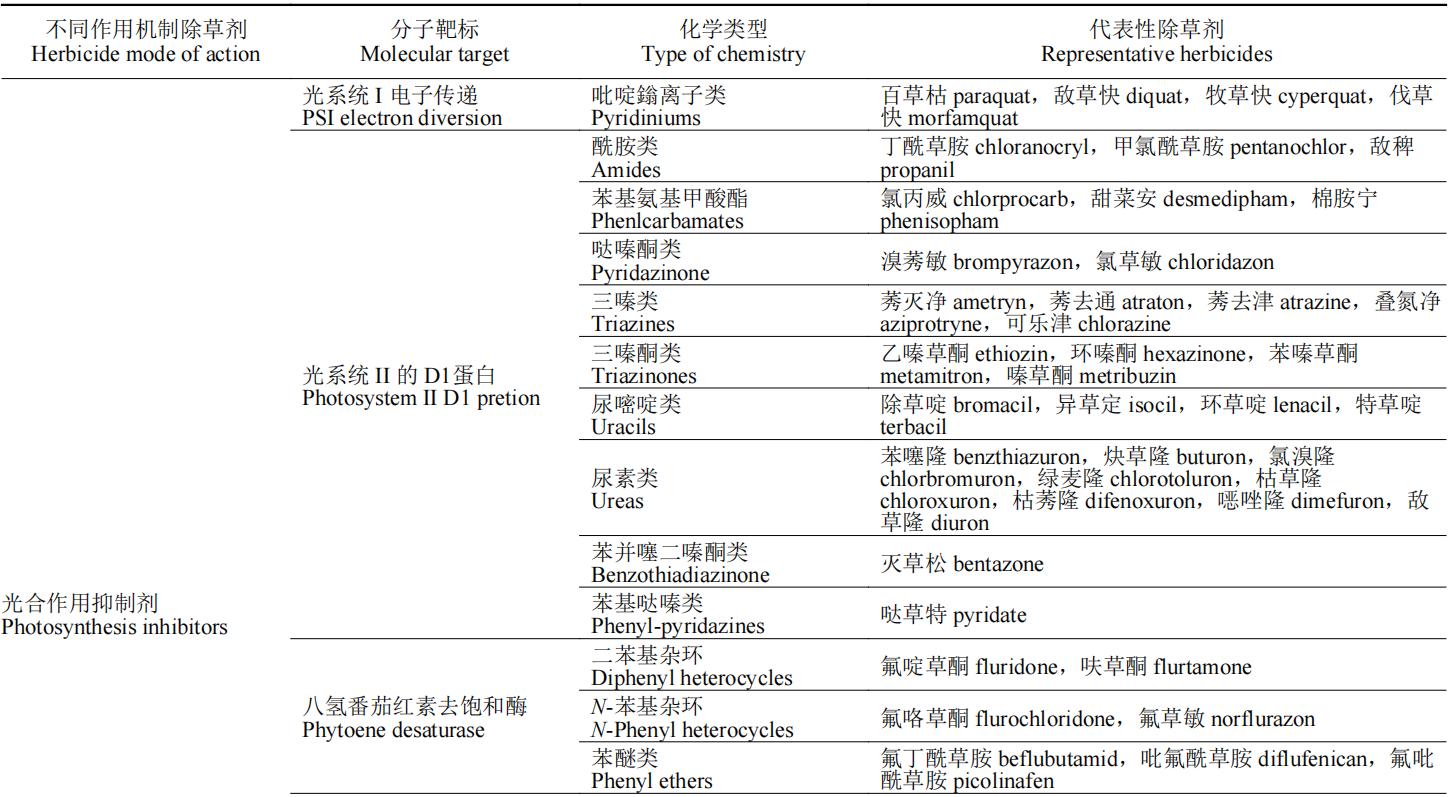
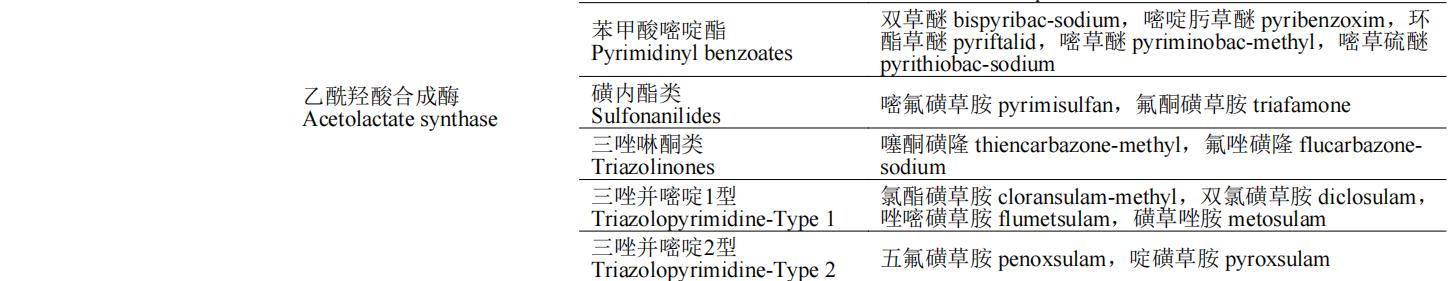
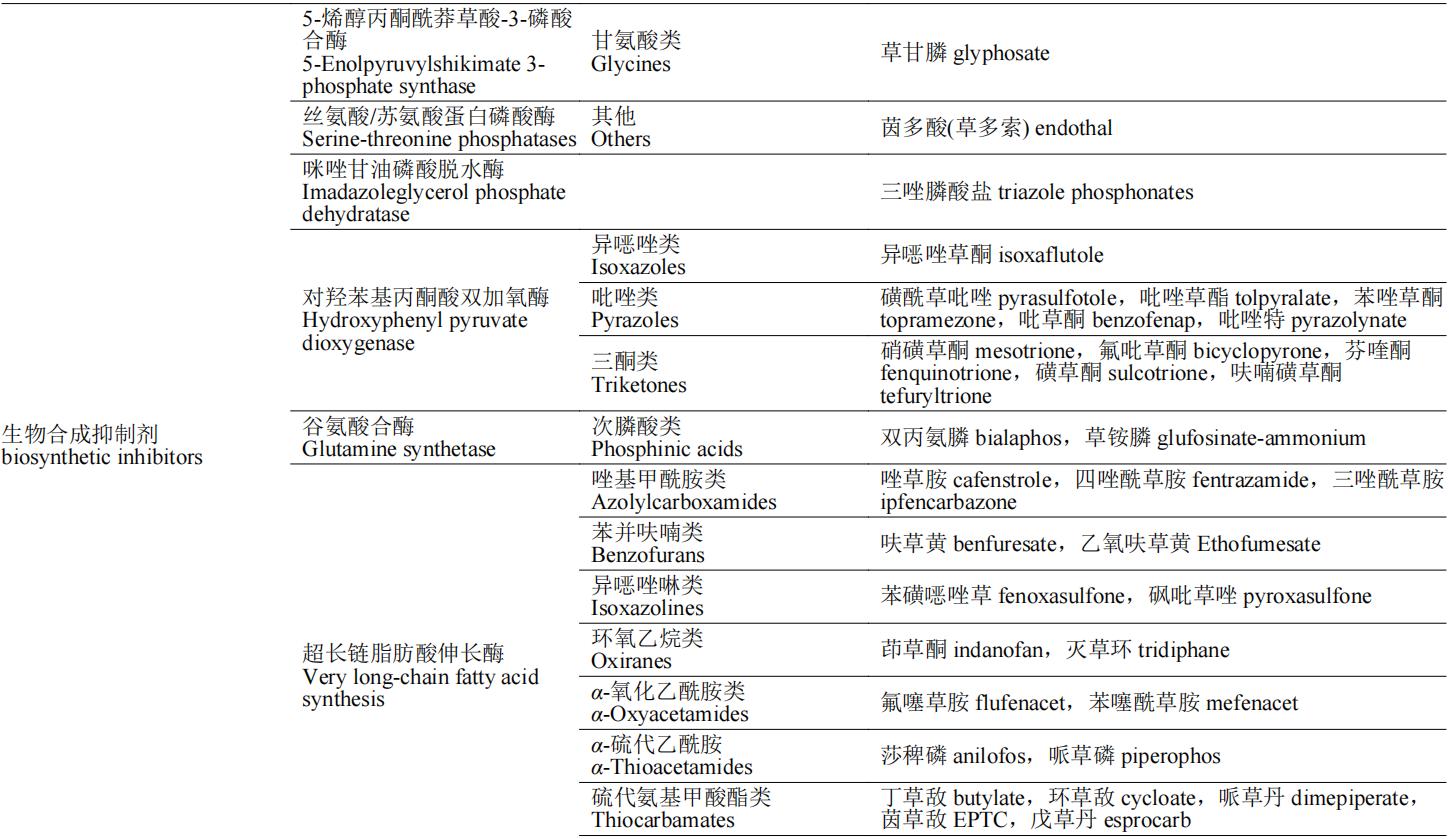
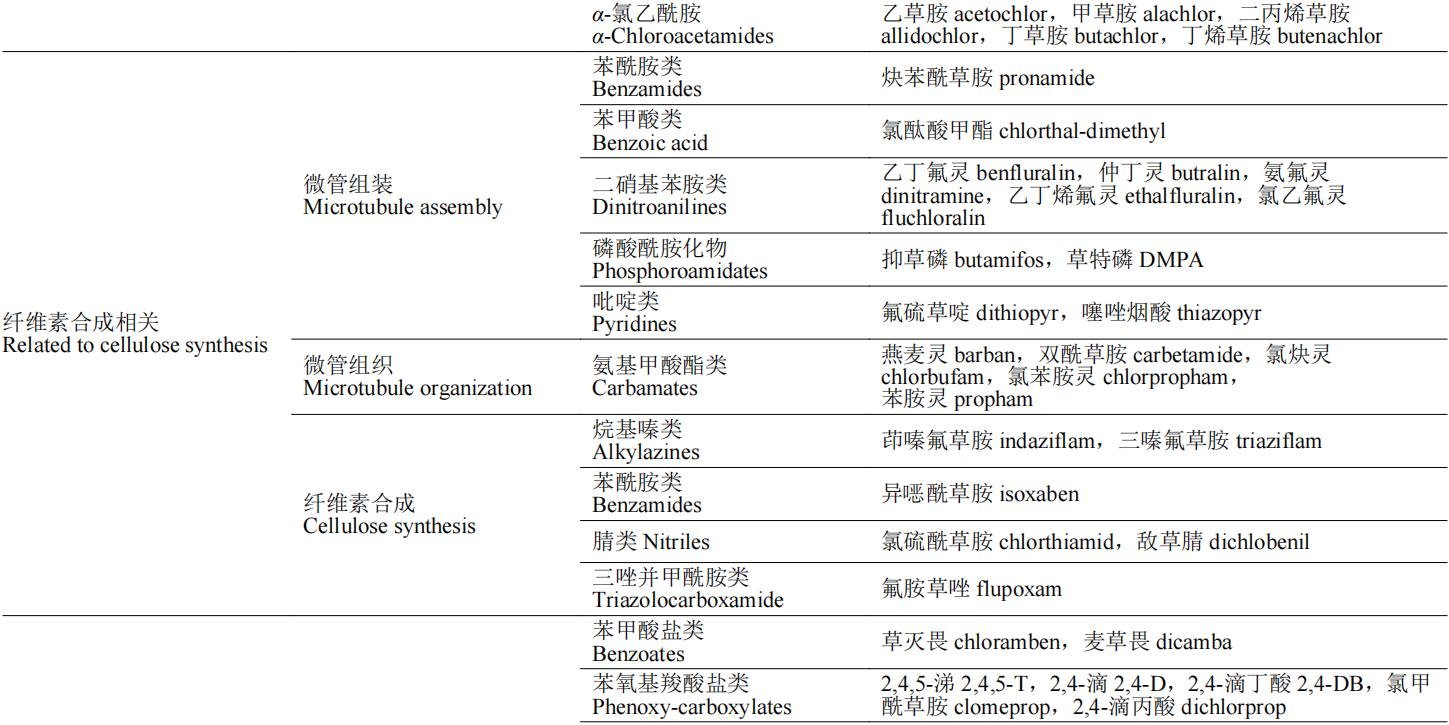
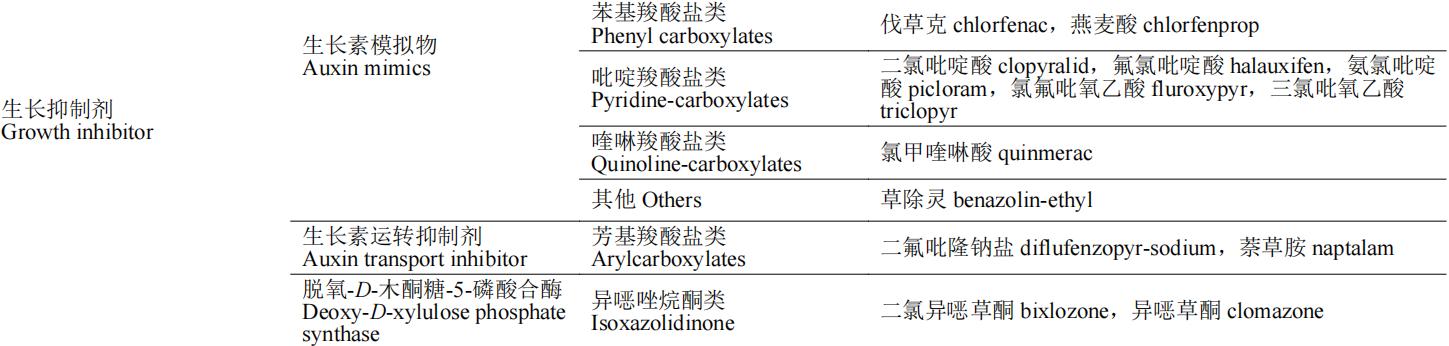
除草剂的作用靶标多为植物生命活动代谢途中的各种酶,如解偶联蛋白是二磷酸腺苷(ADP)生成三磷酸腺苷(ATP)氧化磷酸化过程的催化酶,原卟啉原氧化酶催化亚铁血红素和叶绿素的合成,这些酶均为植物生长必需酶,若缺失或阻断这些酶的合成途径,将会导致植物出现不同症状的死亡。按照除草剂作用机制的不同,可将除草剂分为:光合作用抑制剂、呼吸作用抑制剂、生物合成抑制剂、纤维素合成相关、生长抑制剂、核苷酸合成相关和其他7类(表1)。本文重点介绍除草剂作用机制对应的分子靶标的生理功能、除草剂的作用机制和代表性品种(防治对象和应用范围)。
表1 除草剂靶标的分类及其代表性除草剂品种


2 除草剂分子靶标的生理功能和三维结构研究
本节拟从除草剂施用后,影响杂草光合作用、呼吸作用、生物合成、纤维素合成、生长抑制、核苷酸合成相关和其他7类出发,详细介绍除草剂分子靶标的生理功能、除草剂施用对象及施用范围,为了解现有除草剂分子靶标以及同一分子靶标不同种属蛋白空间活性的差异性,对每个靶标不同种属的蛋白制作了相应的A链三维叠合图。
2.1 光合作用抑制剂
2.1.1 光系统I电子传递
光合作用是绿色植物(包括某些细菌)将光能转化为自身所需的糖类等有机物的生化过程。光合作用又分为两个过程:光系统Ⅰ和光系统Ⅱ。光系统Ⅰ较多分布于类囊体膜基质膜区,是由蛋白亚基组成的色素蛋白复合物,其主要作用为:在光合电子传递链中,将电子从类囊体膜内转移到膜外铁氧还蛋白,在铁氧还蛋白——辅酶Ⅱ(NADP)还原酶作用下,将NADP+还原为还原型辅酶Ⅱ(NADPH)。抑制光系统Ⅰ的除草剂是阻断植物的光电子传递,破坏还原型辅酶Ⅱ的形成,进而阻断植物进行光合作用。百草枯(paraquat)是这类除草剂的代表,其作用机理为:在一定条件下,植物光合膜产生的电子与百草枯离子反应生成自由基,该自由基被植物体内的氧气还原,产生活性氧(ROS),高活性ROS不仅能攻击细胞膜的不饱和脂肪酸产生脂自由基,还能迅速打开并分解细胞膜及组织,导致植物体内细胞组织严重失水,最后植物因失水过多而枯萎死亡。
2.1.2 光系统Ⅱ的D1蛋白
光系统Ⅱ是植物将电子从水中转移到质体醌中,质体醌再与D1蛋白结合。D1蛋白是光系统Ⅱ反应中心一种重要的亚基蛋白,该蛋白可为各种辅助因子提供结合位点,与原初电荷分离和传递有关,如果D1蛋白被破坏,会导致光合电子传递受阻进而影响光系统Ⅱ反应中心,而影响光合作用。抑制光系统Ⅱ的除草剂是一类与质体醌竞争D1蛋白的药剂,通过与D1蛋白结合,阻断电子从QA位点传到QB位点,抑制光合作用的发生。莠去津(atrazine)是这类除草剂的代表,主要用于玉米、甘蔗、高粱等地的芽前、芽后化学除草。施用莠去津后,杂草通过根部吸收莠去津并向上传导,莠去津会与质体醌竞争D1蛋白。当莠去津与D1蛋白结合后,将导致质体醌与D1蛋白的结合和电子传递受阻,这些电子就会与细胞膜中的油脂反应,破坏细胞膜,最终导致杂草死亡。
2.1.3 八氢番茄红素去饱和酶
类胡萝卜素参与植物光合作用,其是类囊体膜上光合元件关键结构的组成部分。八氢番茄红素去饱和酶(PDS)是类胡萝卜素合成途径中的一个限速酶,八氢番茄红素通过PDS两步脱氢,形成ζ-胡萝卜素,ζ-胡萝卜素在ζ-胡萝卜素脱氢酶的作用下,最终形成番茄红素。呋草酮(flurtamone)是这类除草剂的代表,其通过抑制杂草中的PDS,阻碍类胡萝卜素的合成而达到除草效果。
本文选取了菠萝泛菌Pantoea ananatis(PDB编号:4DGK,分辨率:2.35 Å (1 Å=0.1 nm))和水稻Oryza sativa L.(PDB编号:5MOG,分辨率:2.77 Å)PDSA链制作蛋白结构叠合图(图2),运用pymol软件叠合,得出均方根偏差(RMSD)约为10.034 Å,水稻蛋白(蓝色)结构和菠萝泛菌蛋白(绿色)结构相同点:两者都有3个结构域,蛋白折叠相似;其不同点:菠萝泛菌的活性空腔比水稻的略大。
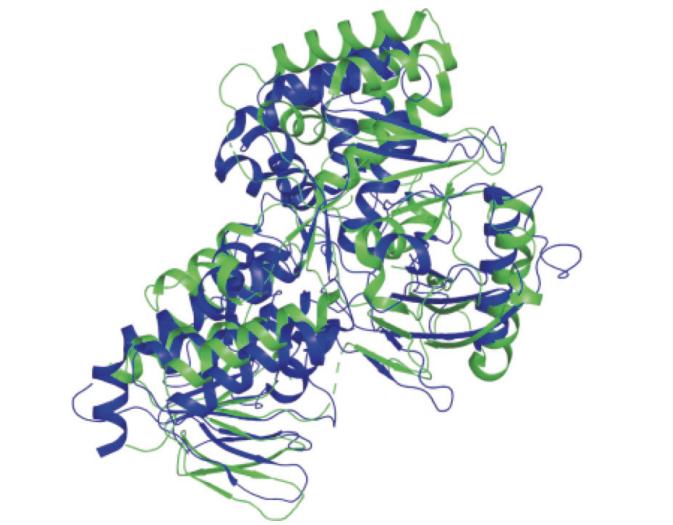
图2 水稻和菠萝泛菌蛋白A链三维结构叠合图
2.1.4 番茄红素环化酶
番茄红素环化酶包括番茄红素β-环化酶和番茄红素ε-环化酶,二者都是类胡萝卜素生物合成途径的酶。抑制番茄红素环化酶的除草剂代表有杀草强(amitrole),该除草剂用于控制一年生杂草和水草(但不能用于粮食作物田杂草的防除)。
2.1.5 原卟啉原氧化酶
原卟啉原氧化酶(PPO)是植物体内四吡咯生物合成途径中最后一个酶。PPO通过催化原卟啉原Ⅸ生成原卟啉Ⅸ,原卟啉Ⅸ是叶绿素和血红素合成途径中物质。抑制PPO的除草剂主要作用机制是:除草剂抑制PPO,造成原卟啉原Ⅸ的积累,导致原卟啉原Ⅸ外泄到细胞质中,在细胞质中迅速被氧化成原卟啉Ⅸ,而细胞质中的原卟啉Ⅸ不能被利用,导致植物光合作用受到影响。抑制PPO的除草剂代表有双唑草腈(pyraclonil),双唑草腈是触杀型除草剂,通过抑制PPO发挥药效作用,能有效抑制各类杂草,如稗草、阔叶杂草和莎草以及耐磺酰脲类除草剂的杂草等。
本文选取了枯草芽孢杆菌Bacillus subtilis(PDB编号:3I6D,分辨率:2.90 Å)、烟草Nicotiana tabacum L.(PDB编号:1SEZ,分辨率:2.90 Å)、黄色粘球菌Myxococcus xanthus(PDB编号:2IVD,分辨率:2.30 Å)和人Homo sapiens(PDB编号:3NKS,分辨率:1.90 Å)的PPO A链制作蛋白结构叠合图,见图3。由图3可知枯草芽孢杆菌蛋白(紫色)和烟草蛋白(蓝色)叠合程度较好,黄色粘球菌(绿色)、人(棕黄色)的蛋白结构与枯草芽孢杆菌和烟草蛋白相似度不高,但两者活性空腔略大。
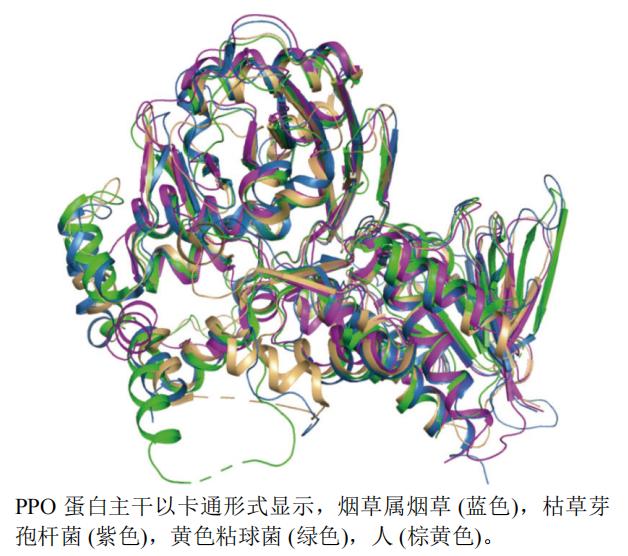
图3 PPO不同种属A链三维结构叠合
2.2 呼吸作用抑制剂——解偶联蛋白
解偶联是指生物呼吸链与氧化磷酸化的偶联遭到破坏的现象。解偶联剂(抑制解偶联)作用于植物氧化磷酸化部位后,由ADP生成ATP的反应受到抑制,导致ADP浓度升高,虽然可以增强植物呼吸作用,却因不能生成ATP来满足植物生长发育的需求,最终导致植物代谢过程损坏。如草不隆(neburon)和环草隆(siduron)是马铃薯块茎线粒体中氧化磷酸化的解偶联剂。
2.3 生物合成抑制剂
2.3.1 乙酰辅酶A羧化酶
植物脂肪酸是油脂以及磷脂的重要组成部分,是重要的能源物质以及信号传导分子。乙酰辅酶A羧化酶(ACCase)催化丙二酰辅酶A的合成,丙二酰辅酶A在质体中用于脂肪酸合成。氰氟草酯(cyhalofop-butyl)是芳氧基苯氧基丙酸酯中唯一对水稻安全性高的除草剂,通过抑制杂草ACCase的活性,使脂肪酸合成停止,细胞生长和分裂不能正常进行,最终导致杂草死亡,此外氰氟草酯对中华钩端草也有较好的防治效果。
本文选取了大肠杆菌Escherichia coli(PDB编号:2F9Y,分辨率:3.20 Å)、嗜热毛壳菌变种DSM 1495(Chaetomium thermophilum var. thermophilum DSM 1495)(PDB编号:5I6I,分辨率:8.40 Å)和酿酒酵母Saccharomyces cerevisiae(PDB编号:5CSA,分辨率:3.00 Å)的ACCase A链制作蛋白结构叠合图(图4),运用pymol软件叠合,从图中可以看出这3类蛋白相似度不高,嗜热毛壳菌变种DSM1495(粉红色)和酿酒酵母蛋白(蓝色)结构比大肠杆菌蛋白(绿色)结构大。
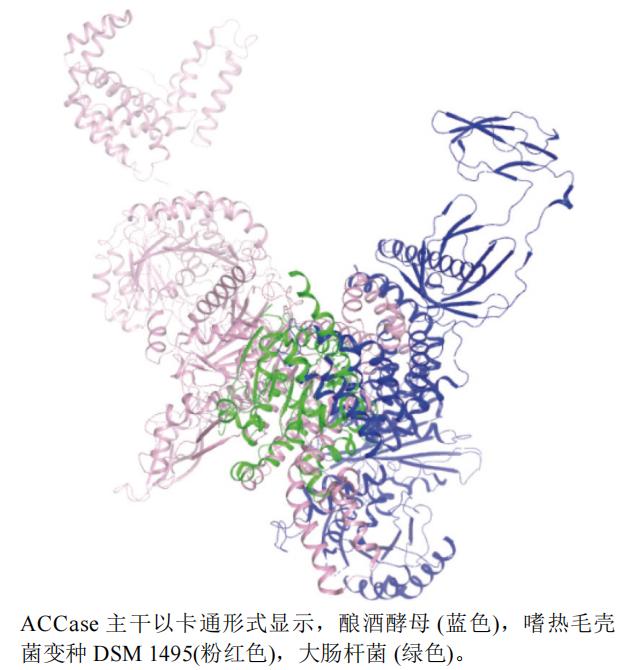
图4 ACCase不同种属A链三维叠合图
2.3.2 脂肪酸硫酯酶
脂肪酸硫酯酶(FAT)是脂肪酸合成过程中控制链长,促使脂肪酸从酰基载体蛋白释放的酶。抑制FAT的除草剂会通过与FAT结合,抑制脂肪酸的释放。环庚草醚(cinmethylin)可抑制FAT活性,作为苗前除草剂,已被引入澳大利亚谷物种植,用于控制一年生黑麦草。甲硫唑草啉(methiozolin)是一种新型除草剂,用于控制几种暖季和冷季草坪草中的一年生蓝草。
2.3.3 茄尼基焦磷酸合酶
质体醌脂质尾的组成部分是二磷酸茄醇,它是通过茄尼基焦磷酸合酶(SPS)的活性获得的,SPS催化七异戊烯基二磷酸依次加成形成香叶基二磷酸。SPS是2020年被Kahlau等证实为除草剂苯草醚(aclonifen)的作用靶标,是一种全新的除草剂作用模式。SPS被抑制,能引起杂草体内质体醌水平急剧下降,最终导致杂草被漂白。
2.3.4 尿黑酸茄尼酯转移酶
尿黑酸茄尼酯转移酶(HST)是质体醌生物合成途径中对羟基苯基丙酮酸双加氧酶的下游酶。HST催化尿黑酸的异戊烯化和脱羧形成2-甲基-6-茄烷基-1,4苯并喹醇,这是质体醌生物合成中的第一个中间体。Shin等在2018年证实,除草剂环哒嗪草酯(cyclopyrimorate)的靶位点是HST,HST被认为是一种新型的商业除草剂分子靶标。
2.3.5 乙酰羟酸合成酶
乙酰乳酸合酶,也称为乙酰羟基酸合酶(AHAS)。AHAS是支链氨基酸生物合成途径中的一个关键酶,支链氨基酸包括亮氨酸、异亮氨酸和缬氨酸,若植物缺乏这3种氨基酸,将影响蛋白质合成,抑制细胞分裂,导致杂草组织失绿、黄化、生长受阻,最后杂草逐渐死亡。除草剂五氟磺草胺(penoxsulam)作用靶点是AHAS,为苗后除草剂,用于水稻田杂草防治。五氟磺草胺可与杂草中AHAS的结合部位竞争性结合,从而影响杂草体内氨基酸的合成,抑制杂草生长,杂草生长点因失绿而逐渐坏死。
本文选取了哈茨木霉Trichoderma harzianum(PDB编号:7EGV,分辨率:2.54 Å)、肺炎克雷伯氏菌Klebsiella pneumoniae(PDB编号:1OZG,分辨:2.30 Å)、海栖热袍菌MSB8 Thermotoga maritima MSB8(PDB编号:2FGC,分辨率:2.30 Å)和枯草芽孢杆菌B. subtilis(PDB编号:4RJI,分辨率:3.20 Å)的AHASA链]制作蛋白结构叠合图,见图5。蛋白叠合图以枯草芽孢杆菌蛋白(蓝色)为主体进行叠合,可以看出,除了海栖热袍菌蛋白(绿色)结构与其他不同,其余蛋白结构相似。
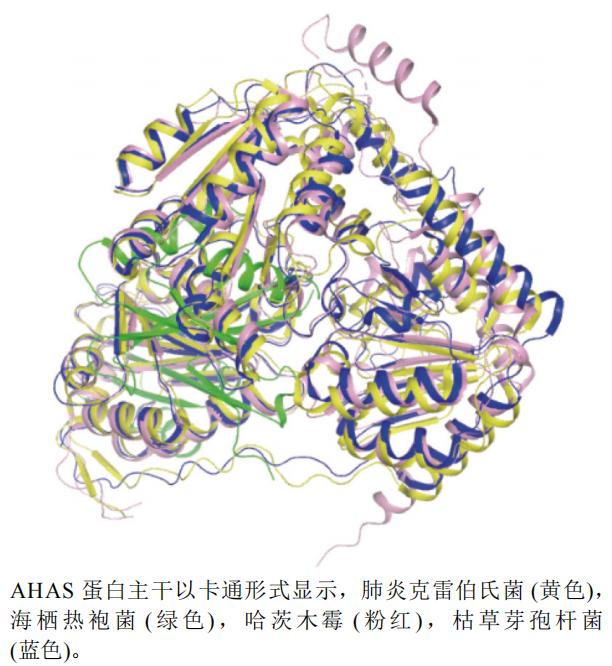
图5 AHAS不同种属A链三维叠合图
2.3.6 二羟基酸脱水酶
二羟基酸脱水酶(DHAD)是支链氨基酸生物合成途径中一种常见酶,在不同的植物物种中高度保守。研究证明,DHAD是一个良好的除草剂分子靶标,作用于DHAD的除草剂不会对哺乳动物造成影响,因为哺乳动物体内并不存在支链氨基酸生物合成途径。加州大学洛杉矶分校一个课题小组使用抗性基因为导向的基因组挖掘技术,发现了天然产物天冬氨酸(aspterric acid)可以靶向作用于DHAD,从而起到抑制植物生长的作用。
本文选取了集胞藻Synechocystis sp.(PDB编号:6NTE,分辨率:2.33 Å)和拟南芥Arabidopsis thaliana(PDB编号:5YMO,分辨率:1.84 Å)的DHADA链制作蛋白结构叠合图(图6),得出RMSD约为1.123 Å,表明两者蛋白结构相似,但集胞藻(绿色)的活性空腔比拟南芥(蓝色)的活性空腔大。
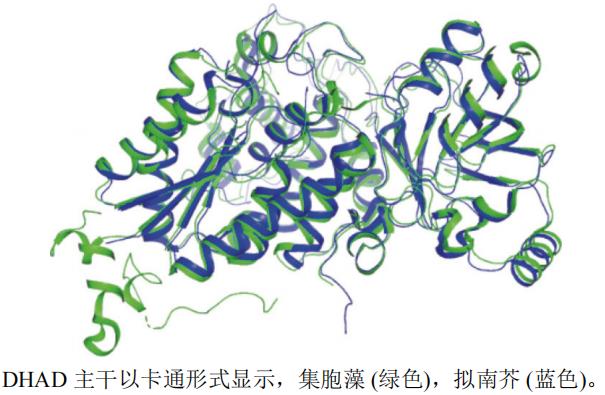
图6 DHAD不同种属A链三维叠合图
2.3.7 5-烯醇丙酮酰莽草酸-3-磷酸合酶
5-烯醇丙酮酰莽草酸-3-磷酸合酶(EPSPS)是叶绿体中的一种酶,它催化莽草酸合成途径的倒数第2步反应,是合成芳香族氨基酸以及部分次生代谢物过程的关键酶。抑制EPSPS,从而抑制莽草素向苯丙氨酸、酪氨酸及色氨酸的转化,使蛋白质合成受到干扰,导致植物死亡。草甘膦(glyphosate)作用于杂草分子靶标的EPSPS,杂草通过茎叶吸收后传导到植物各部位,可防除单子叶、双子叶、一年生、多年生、草本和灌木等40多科植物。
本文选取了大肠杆菌E. coli(PDB编号:1MI4,分辨率:1.70 Å)、肺炎链球菌Streptococcus pneumoniae(PDB编号:1RF4,分辨率:2.20 Å)和白假丝酵母Candida albicans(PDB编号:7TBU,分辨率:1.85 Å)EPSPSA链制作蛋白结构叠合图(图7)。运用pymol软件以大肠杆菌蛋白(绿色)结构为主体进行叠合后发现,白假丝酵母蛋白(粉红色)结构活性空腔比大肠杆菌和肺炎链球菌蛋白(蓝色)活性空腔大。
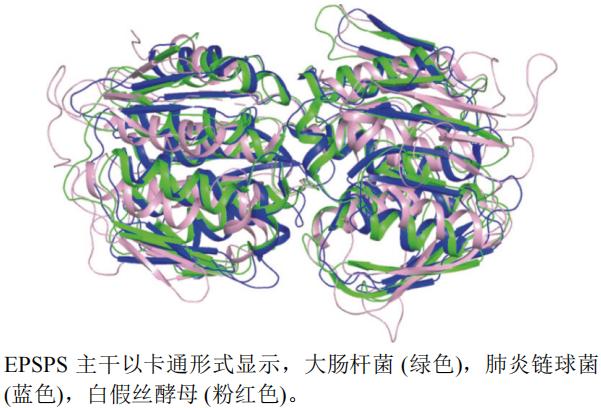
图7 EPSPS不同种属A链三维叠合图
2.3.8 丝氨酸-苏氨酸蛋白磷酸酶
丝氨酸-苏氨酸蛋白磷酸酶催化结构域在动物、植物和原生动物中高度保守。但张晓晨等认为丝氨酸/苏氨酸磷酸酶是斑蝥素(水泡甲虫和西班牙苍蝇的天然产物)及其类似物作用于木霉菌的靶标。丝氨酸-苏氨酸蛋白磷酸酶是茵多酸(endotha)的靶标,茵多酸诱导严重的生长抑制,用于管理水生环境中的杂草。
本文选取了酿酒酵母Saccharomyces cerevisiae(PDB编号:3ICF,分辨率:2.30 Å)、人H. sapiens(PDB编号:3H67,分辨率:1.65 Å)、白假丝酵母Candida albicans(PDB编号:5JPE,分辨率:2.61 Å)、埃希氏菌病毒λ(Escherichia virus lambda)(PDB编号:1G5B,分辨率:2.15 Å)、褐家鼠Rattus norvegicus(PDB编号:4JA7,分辨率:2.00 Å)和拟南芥A. thaliana(PDB编号:5JJT,分辨率:2.10 Å)的丝氨酸-苏氨酸蛋白磷酸酶A链制作蛋白结构叠合图(图8)。运用pymol软件,以人(蓝色)的蛋白结构为主体叠合后发现,埃希氏菌病毒λ(绿色)和拟南芥(墨绿色)的蛋白结构活性空腔较其他蛋白活性空腔大。
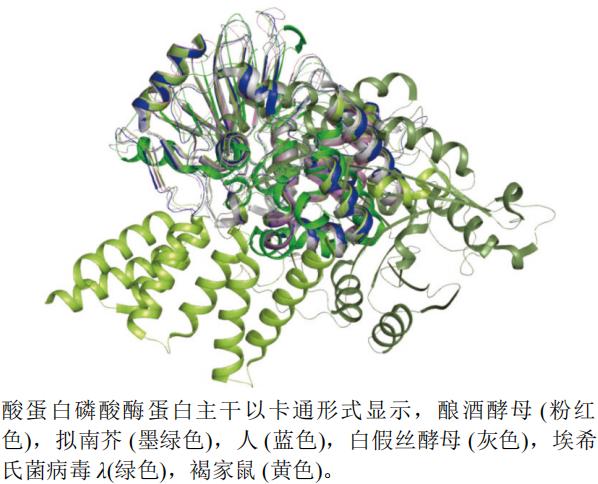
图8 丝氨酸-苏氨酸蛋白磷酸酶不同种属A链三维叠合图
2.3.9 咪唑甘油磷酸脱水酶
咪唑甘油磷酸脱水酶也是一种良好的除草剂靶标,动物体内不存在咪唑甘油磷酸脱水酶。咪唑甘油磷酸脱水酶催化植物和微生物中组氨酸生物合成的重要步骤,在组氨酸生物合成途径中催化咪唑甘油磷酸酯转化为咪唑乙酰醇磷酸酯。已经证实咪唑甘油磷酸脱水酶抑制剂具有广谱苗后除草活性,3种三唑磷酸显著抑制咪唑甘油磷酸脱水酶的活性,细胞培养中显示很强的抑制作用。
2.3.10 对羟苯基丙酮酸双加氧酶
对羟苯基丙酮酸双加氧酶(HPPD)是生物体内酪氨酸代谢过程中重要的酶,酪氨酸在酪氨酸氨基转移酶的作用下生成对羟基苯丙酮酸,在氧气的参与下HPPD能够将对羟基苯丙酮酸催化转化成尿黑酸。在植物体内,尿黑酸能够被进一步转化成质体醌和生育酚。当HPPD的活性被抑制,杂草体内酪氨酸正常代谢将被阻断,导致杂草体内类胡萝卜素的缺乏,从而使叶绿素光氧化作用减弱,影响植物的光合作用,进而使杂草白化而死亡。硝磺草酮(mesotrione)为独特的玉米田除草剂,对玉米田一年生阔叶杂草和部分禾本科杂草(如苘麻、苋菜、藜、蓼、稗草、马唐等)有较好的防治效果,而对铁苋菜和一些禾本科杂草防治效果较差。
本文选取了玉米Zea mays L.(PDB编号:1SP8,分辨率:2.00 Å)、荧光假单胞菌Pseudomonas fluorescens(PDB编号:1CJX,分辨率:2.40 Å)、阿维链霉菌Streptomyces avermitilis(PDB编号:1T47,分辨率:2.50 Å)、人H. sapiens(PDB编号:6ISD,分辨率:2.40 Å)和拟南芥A. thaliana(PDB编号:1SP9,分辨率:2.13 Å)的HPPDA链制作蛋白叠合图(图9)。运用pymol软件以荧光假单胞菌蛋白(绿色)为主体进行叠合后发现,以上5个蛋白中有一处叠合情况不好。
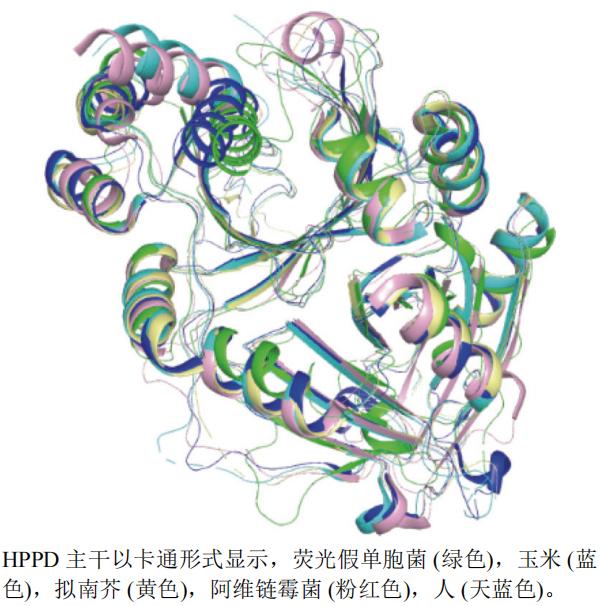
图9 HPPD不同种属A链三维叠合图
2.3.11 谷氨酰胺合成酶
谷氨酰胺合成酶(GS)在微生物及植物体中参与谷氨酰胺合成和氮的循环。谷氨酰胺合成是高等植物氮同化的第一步,在GS催化下,氨掺入谷氨酰胺。氨参杂的产生的谷氨酰胺易于代谢,为后续含氮化合物生物合成的提供基础。草铵膦(glufosinate-ammonium)以GS为靶标,通过抑制GS的活性,造成杂草体内氮代谢紊乱和氨过量积累,导致叶绿体解体,破坏杂草光合作用,达到除草效果。
本文选取了玉米Z. mays(PDB编号:2D3A,分辨率:2.63 Å)、青春双歧杆菌Bifidobacterium adolescentis(PDB编号:4S17,分辨率:2.30 Å)、结核分枝杆菌Mycobacterium tuberculosis(PDB编号:1HTO,分辨率:2.40 Å)、人H. sapiens(PDB编号:2OJW,分辨率:2.05 Å)的GSA链制作蛋白结构叠合图(图10)。运用pymol软件以结核分枝杆菌蛋白(绿色)为主体进行叠合后发现,以上4个蛋白叠合程度并不是很好,一些结构域也没有相似性。
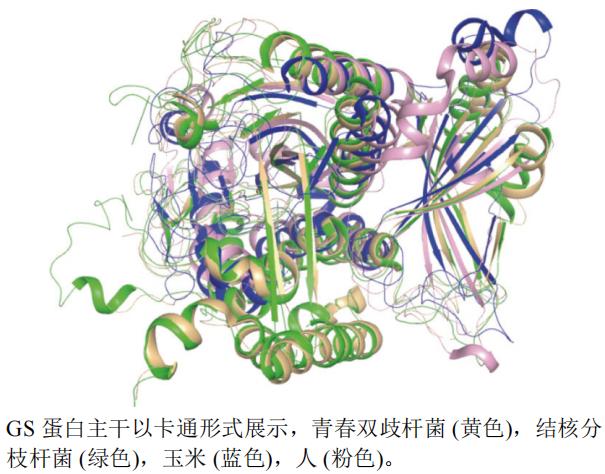
图10 GS不同种属A链三维叠合图
2.3.12 超长链脂肪酸合酶
超长链脂肪酸合酶在代谢途径负责合成蜡、角质素和木栓素。砜吡草唑(pyroxasulfone)属超长链脂肪酸合成酶抑制剂除草剂,主要作为芽前封闭处理剂,以封杀禾本科为主、阔叶草为辅,具有诸多优势。适用作物范围广,可以用于小麦、玉米、花生、水稻、大豆和棉花等作物。
2.4 纤维素合成相关
2.4.1 微管组装
植物微管具有重要的生物学功能,如维持植物细胞形态结构、参与细胞运动、细胞内物质运输、信号传导、细胞分裂等,可参与形成纺锤体、基粒、中心粒、轴突、神经管、纤毛、鞭毛等结构。抗微管类除草剂是通过作用于细胞微管而影响纺锤体形成,并抑制细胞有丝分裂。二硝基苯胺类除草剂是抑制微管的典型代表,可与微管蛋白结合并抑制其聚合,造成纺锤体微管丧失,使杂草细胞有丝分裂停留于前期或中期,产生异常的多形核。由于杂草细胞极性丧失,液泡形成能力增强,故在伸长区进行放射性膨胀,结果造成根尖肿胀。
2.4.2 微管组织
微管组织中心是真核细胞中形成微管的结构,有两个主要的功能:1)组织真核细胞中鞭毛和纤毛的形成;2)组织真核细胞减数分裂或有丝分裂过程中纺锤体的形成。抑制微管组织的除草剂通常为氨基甲酸酯类,有燕麦灵(barban)和氯苯胺灵(chlorpropham),它们通过破坏杂草微管的稳定性来破坏有丝分裂而达到除草作用。
2.4.3 纤维素合成
植物纤维是纤维素与各种营养物质结合生成的丝状或絮状物,对植物具有支撑、连接、包裹和充填等作用。茚嗪氟草胺(indaziflam)为抑制纤维素合成的除草剂,可抑制各向异性细胞扩增,导致严重肿胀和发育迟缓的生长表型。
2.5 生长抑制剂
2.5.1 生长素模拟物
植物生长素是由具有分裂和增大活性的细胞区产生调控植物生长速度和方向的激素。生长素使植物细胞壁松弛,从而使细胞生长伸长,在许多植物中还能增加核糖核酸和蛋白质的合成。因生长素的两重性,生长模拟物对某类型杂草具有杀害作用。如2,4-滴(2,4-D),它是一种选择性除草剂,阔叶除草剂,在不影响单子叶植物的情况下杀死双子叶植物,并在分子水平上模仿天然生长素。
2.5.2 生长素转运抑制剂
生长素转运是植物生长和发育过程所必需的,包括重力影响和侧根生长。TIBA(2,3,5-triiodobenzoic acid)和NPA(naphthylphthalamic acid)是两种最常用的生长素转运除草剂,其在杂草生长过程中阻碍生长素的转运,进而杀死杂草。
2.5.3 脱氧-D-木酮糖-5-磷酸合酶
脱氧-D-木酮糖-5-磷酸合成酶(DXS)是甲羟戊酸(MEP)途径的第一个限速酶,也是该途径的关键调控酶,对单萜和双萜类香气化合物以及类胡萝卜素、叶绿素等重要物质的合成具有关键调控作用。抑制DXS的除草剂有二氯异噁草酮(bixlozone)和异噁草酮(clomazone)。
本文选取了大肠杆菌E. coli(PDB编号:2O1S,分辨率:2.40 Å)、耐辐射奇球菌Deinococcus radiodurans(PDB编号:2O1X,分辨率:2.90 Å)和拟南芥A. thaliana(PDB编号:7BZX,分辨率:4.00 Å)的DXSA链制作蛋白结构叠合图(图11)。运用pymol软件以拟南芥(粉色)为主体叠合后发现,三者蛋白结构相似,仅有一处蛋白不相似。
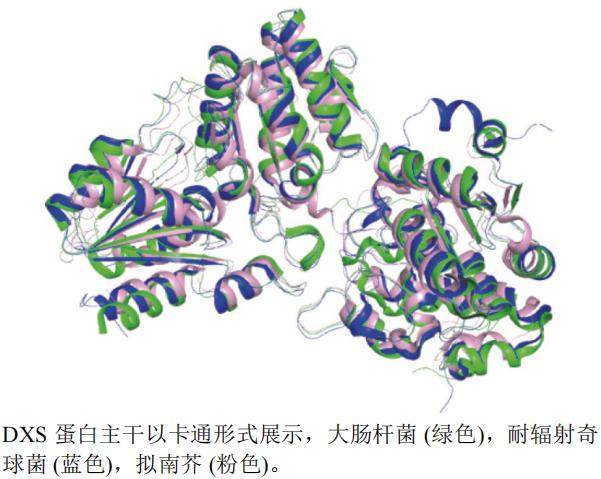
图11 DXS不同种属A链三维叠合图
2.6 其他
2.6.1 二氢乳清酸脱氢酶
嘧啶从头生物合成由6个酶促步骤组成,其中第4步由二氢乳清酸脱氢酶(DHODH)催化。该酶将泛醌介导的二氢乳清酸氧化为乳清酸,所有杂草DHODH都是位于线粒体内膜外表面的黄素蛋白,由于核苷酸的核心作用,该途径的抑制对大多数生物都是致命的。目前开发的通用化学名称被临时批准为tetflupyrolimet的除草剂对水稻田的杂草控制具有选择性。tetflupyrolimet竞争DHODH上的泛醌结合位点,用tetflupyrolimet处理的敏感植物没有萎黄,但会发展出独特的发育迟缓表型,表明它们缺乏生长的关键分子(嘧啶)。
本文选取了人H. sapiens(PDB编号:1D3G,分辨率:1.60 Å)、黑鼠Rattus rattus(PDB编号:1UUM,分辨率:2.30 Å)、布氏锥虫Trypanosoma brucei(PDB编号:2B4G,分辨率:1.95 Å)和乳酸乳球菌Lactococcus lactis(PDB编号:2BX7,分辨率:2.04 Å)的DHODHA链制作蛋白结构叠合图(图12)。运用pymol软件以布氏锥虫(粉色)为主体进行叠合后发现,人(绿色)和黑鼠(蓝色)的蛋白结构与其他两个蛋白部分结构不相似。
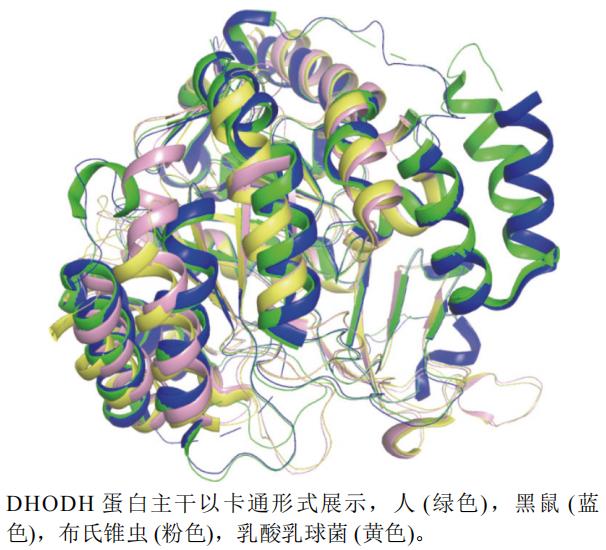
图12 DHODH不同种属A链三维叠合图
2.6.2 二氢蝶酸合成酶
叶酸是植物生长发育所需的重要功能因子,包括四氢叶酸及其衍生物,属于水溶性B族维生素(B9)。叶酸缺乏和叶酸稳态变化会严重影响植物胚胎发育、幼苗生长、开花和结实等过程。抑制叶酸的除草剂黄草灵(asulam)用于棉田、大豆、谷物、甜菜等作物中防除狗尾草、冰草、田蓟、马唐、稗草等。
本文选取了大肠杆菌E. coli(PDB编号:1AJ0,分辨率:2.00 Å)、嗜热栖热菌Thermus thermophilus(PDB编号:2DQW,分辨率:1.65 Å)、肺炎链球菌Streptococcus pneumoniae(PDB编号:2VEF,分辨率:1.80 Å)和新洋葱伯克霍尔德氏菌Burkholderia cenocepacia(PDB编号:2Y5S,分辨率:1.95 Å)的二氢蝶酸合成酶A链制作蛋白结构叠合图(图13)。运用pymol软件以新洋葱伯克霍尔德氏菌(粉色)为主体进行叠合后发现,肺炎链球菌蛋白(黄色)结构活性空腔比其他3个蛋白的大。
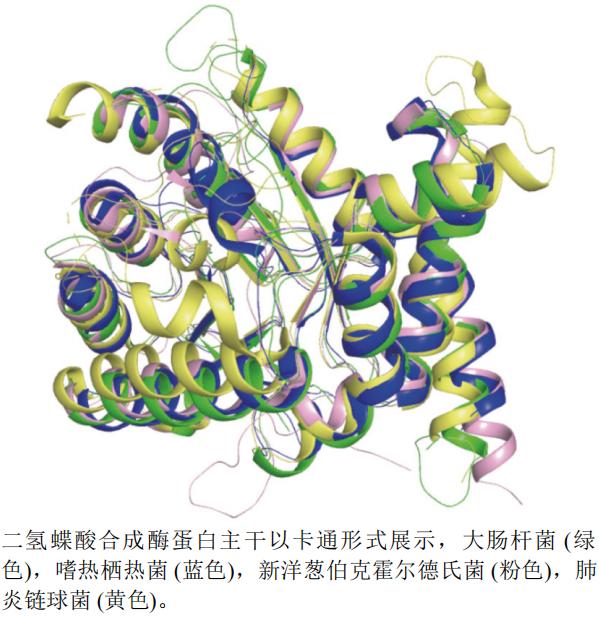
图13 二氢蝶酸合成酶蛋白不同种属A链三维叠合图
3 除草剂分子靶标发现的现状和展望
3.1 除草剂分子靶标发现的现状
目前除草剂分子靶标发现面临很多问题:1)草甘膦和抗草甘膦作物的协同使用在很大程度上减弱了除草剂靶标发现的驱动力。2)除草剂开发公司的合并、减少等,也导致从事此类研究的科学家数量大幅减少,进而减少除草剂靶标发现。3)随着环保要求的提高以及新农药登记制度更加严格,新型除草剂靶标发现的成本也显著增加。一个典型表现是近些年将合成农药推向市场的成本增至2.86亿美元,新除草剂靶标发现和新药创制难度越来越大。
3.2 除草剂分子靶标发现的新技术及展望
除草剂新分子靶标的发现是目前解决除草剂抗性问题最好方法之一。这里介绍几种发现新分子靶标的方法:一是以基因组学为代表的新生物技术,Duke等通过对微生物基因簇编码,来生产倍半萜天蝶酸的生物合成途径和其靶酶抗性酸形式的基因,证明了基因组方法为发现具有新模式作用的除草剂提供了一种新的策略。二是人工智能技术,基于计算机筛选和大型分子数据库的过滤,体外选择分子靶标,确定有可行的新除草剂分子靶标。三是多学科推进发展,如Sukhoverkov等验证了抑制叶绿体的翻译,可作为新除草剂靶标。Hall等发现在氨基酸生物合成中有尚未商业化开发的潜在除草剂靶标。近些年,有一些新作用方式的除草剂被发现:HMGR(3-羟基-3-甲基戊二酰辅酶A还原酶)是真核生物甲羟戊酸途径上的限速酶,用于治疗人类高胆固醇血症,Haywood等证明,可修饰HMGR抑制性特点,使其有望成为除草剂靶标;科罗拉多州立大学杂草研究实验室报告了关于导致草铵膦接触活性的因素的新见解,发现草铵膦因触发活性氧物种(ROS)快速、大量产生,导致细胞膜灾难性脂质过氧化和细胞快速死亡,其效果与除草剂的吸收成正比。这些发现启发我们可以在原有的靶标上开发新除草剂,或者在植物体生长代谢途中再去寻找新的除草剂靶标位点。
本文描述了近些年除草剂靶标的生理功能、除草剂的作用原理以及适用范围,在此基础上提出了新除草剂靶标发现的困境,也简单介绍了近些年一些新作用方式的除草剂和靶标发现的新技术,可为继续开发新的除草剂靶标提供参考。
(1)本网旨在传播信息,促进交流,多方面了解农药发展动态,但不构成任何投资建议。
(2)所有文章仅代表作者观点,不代表本网立场。
(3)“信息来源:江苏省农药协会 农药资讯网”为原创文章,转载时请注明来源和作者。
(4)本网转载文章及图片的版权属于原作者,若有侵权,请联系删除。