




1962年加拿大尤尼罗亚尔(Uniroyal)公司的Ruprecht申请了氧硫杂芑顺式丁烯酰替苯胺类杀菌剂萎锈灵(carboxin,2,3-二氢-5-(N-甲酰苯胺)-6-甲基-1,4-氧硫杂芑)的专利(US3933847A),并于1966年商品化,用于种子和土壤处理,防治作物黑穗病。萎锈灵对担子菌的专化性、内吸输导性和治疗作用,引起了人们对其作用机制和克服其易被氧化及光解问题的研究。1970年美国蒙大拿州立大学的Mathre发现萎锈灵的作用靶标是呼吸链中的琥珀酸脱氢酶(复合物Ⅱ),后来进一步研究发现,萎锈灵作用于琥珀酸脱氢酶B亚基。经过20多年的研究探索,至20世纪80年代中后期,日本住友化学株式会社、组合化学工业株式会社、日产化学工业株式会社分别开发出呋吡菌胺(furametpyr)、灭锈胺(mepronil)和噻呋酰胺(thifluzamide),并于90年代商品化用于防治水稻纹枯病。此后,各大农药企业进一步基于靶标结构生物学纷纷研发出结构多样、广谱、高效、低毒、持效性长的琥珀酸脱氢酶抑制剂(Succinate dehydrogenase inhibitors,SDHIs),目前已经成为继麦角甾醇生物合成抑制剂类(Ergosterol biosynthesis inhibitors,EBIs)和Qo位点呼吸抑制剂类(Quinone outside inhibitors,QoIs)之后,杀菌剂市场占有率第3大的新型选择性杀菌剂。但是,随着SDHIs杀菌剂的大量使用,多种植物病原真菌已经对该类药剂产生了抗性,且抗性菌株在病原群体中的比例和抗性发生范围不断升高,如果不能及时采取抗药性治理措施,必将影响这类杀菌剂的使用寿命。2010年,Hervé综述了SDHIs杀菌剂的作用机制和抗性进化机制,2013年,Helge综述了SDHIs杀菌剂的抗性研究进展。本文阐述SDHIs杀菌剂的发展史及其作用机制,重点综述植物病原真菌对该类药剂的抗性发生发展、抗性机制及抗性治理策略。
1 SDHIs杀菌剂的开发和分类
SDHIs是作用于细胞线粒体呼吸链复合物Ⅱ的新型杀菌剂,因其化学结构中均含有酰替苯胺活性基团,又称酰替苯胺类杀菌剂,根据酰胺键连接的官能团不同又可细分为吡唑酰胺类、吡啶酰胺类、苯基酰胺类等。SDHIs杀菌剂属线粒体呼吸链抑制剂,因真菌孢子萌发对能量及依赖能量代谢产生的小分子碳水化合物需求旺盛,以致SDHIs杀菌剂对孢子萌发的抑制活性常常显著高于对菌丝生长的抑制活性。新型SDHIs杀菌剂突破了早期开发的SDHIs杀菌剂如萎锈灵(carboxin)等有关生物学和理化性质的局限性,不仅对担子菌表现很高的抗菌活性,而且对多种子囊菌同样表现高活性,且克服了易被氧化光解的缺点,可以喷施防治多种作物病害。自巴斯夫于2003年上市抗菌谱广、活性高、环境风险低、内吸性和传导性好的啶酰菌胺(boscalid)以后,各大农药公司纷纷投入大量精力和财力开展了更加安全、高效、广谱的新一代SDHIs杀菌剂研发,并将其陆续商品化。如吡唑萘菌胺(isopyrazam,2010年,先正达)、联苯吡菌胺(bixafen,2011年,拜耳)、氟唑菌苯胺(penflufen,2012年,拜耳)、异丙噻菌胺(isofetamid,2015年,日本石原)、氟唑菌酰羟胺(pydiflumetofen,2016年,先正达)、联苯吡嗪菌胺(pyraziflumid,2018年,日本农药)等陆续推向杀菌剂市场(表1)。
自然界也存在天然的与SDHIs杀菌剂结构类似的活性物质,如Promysalin是一种Pseudomonad aeruginosa的次生代谢物,具有抗菌活性,最初从植物根际中分离出来,亲和蛋白图谱(affinity-based protein profiling)鉴定表明,琥珀酸脱氢酶为该天然产物的生物学靶点。目前,已经有25个SDHIs类杀菌剂品种进入杀菌剂市场,此类产品已成为杀菌剂市场销售额增长最为迅速的一类药剂,在重要植物病害的化学防控中发挥巨大作用。
现今的研究表明,SDHIs杀菌剂的结构包括三部分:酸片段、胺片段和酰胺键连接部分。结合国际杀菌剂抗性行动委员会(Fungicide Resistance Action Committee,FRAC)2021年公布的SDHIs杀菌剂的化学结构细分归纳于表1。
表1 琥珀酸脱氢酶抑制剂类杀菌剂的化学结构分类
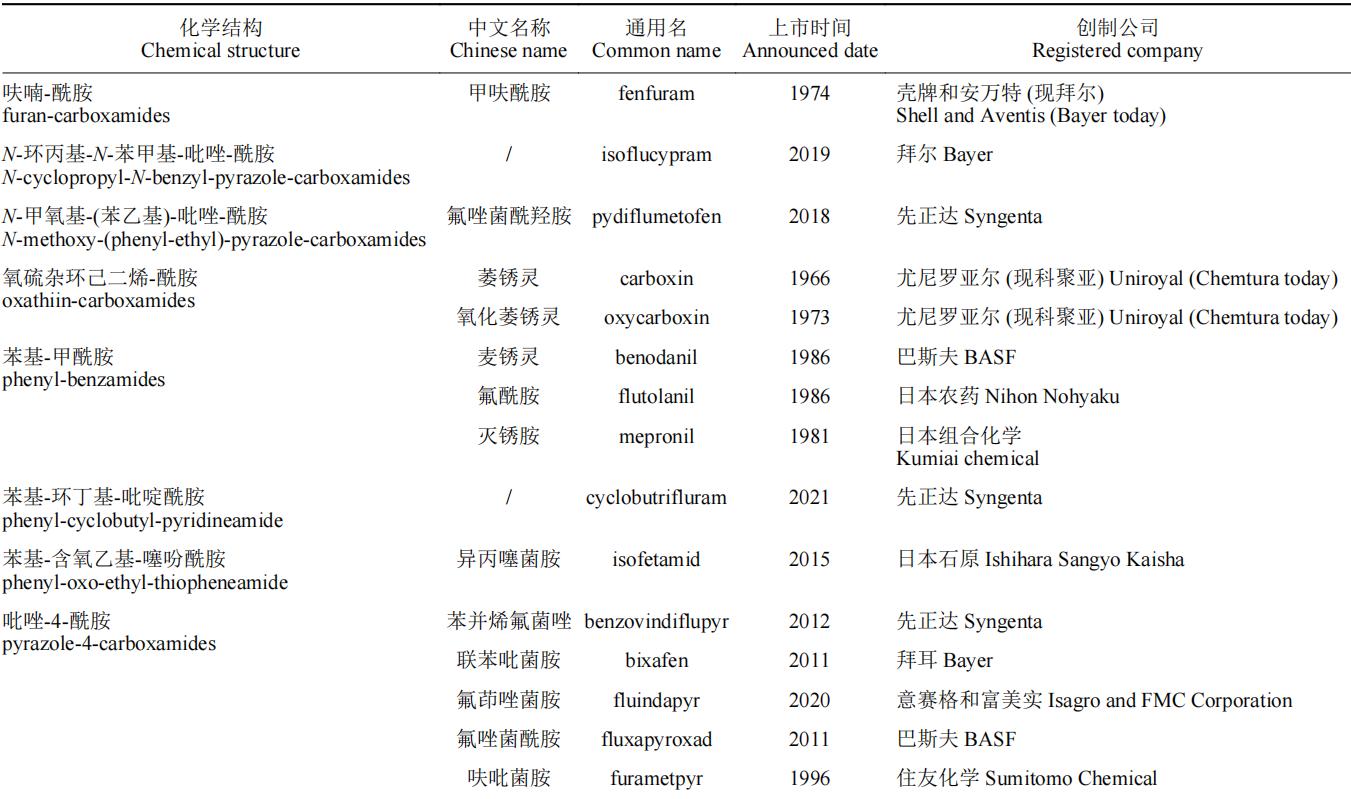
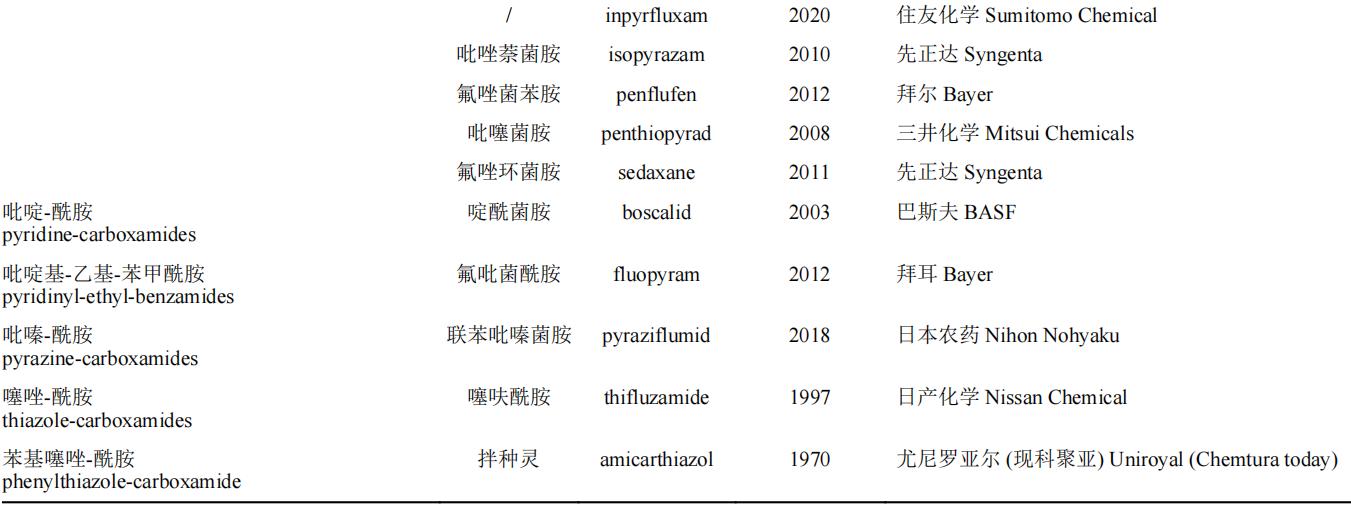
注:“/”代表当前该药剂尚无对应中文名。
在SDHIs的结构中,活性基团为酰胺键(氟唑菌酰羟胺(pydiflumetofen)为羟基酰胺键,氟唑菌酰羟胺在同类药剂中活性极其优异,可能与酰胺键的改造有关),其余两个基团为改造基团。有研究表明,酰胺键与胺片段的距离对药剂的活性有显著影响。此外,苯甲酰胺类杀菌剂的氟吡菌胺(fluopicolide)和我国自主研发的氟醚菌酰胺(fluopimomide)曾被认为是SDHIs杀菌剂,根据最新的研究结果,FRAC将氟吡菌胺和氟醚菌酰胺(fluopimomide)的作用机制归为作用于细胞骨架和马达蛋白的delocalisation of spectrin-like proteins亚类(作用机制编码为B5,抗性编码为#43)。
2 SDHI杀菌剂的应用现状
SDHIs杀菌剂以其广谱性在杀菌剂市场上占据了重要地位。在我国,SDHIs杀菌剂在各类粮食作物和经济作物病害防治上均有登记(中国农药信息网)。部分药剂的单剂登记现状见表2。
表2 SDHIs杀菌剂在中国的病害登记情况
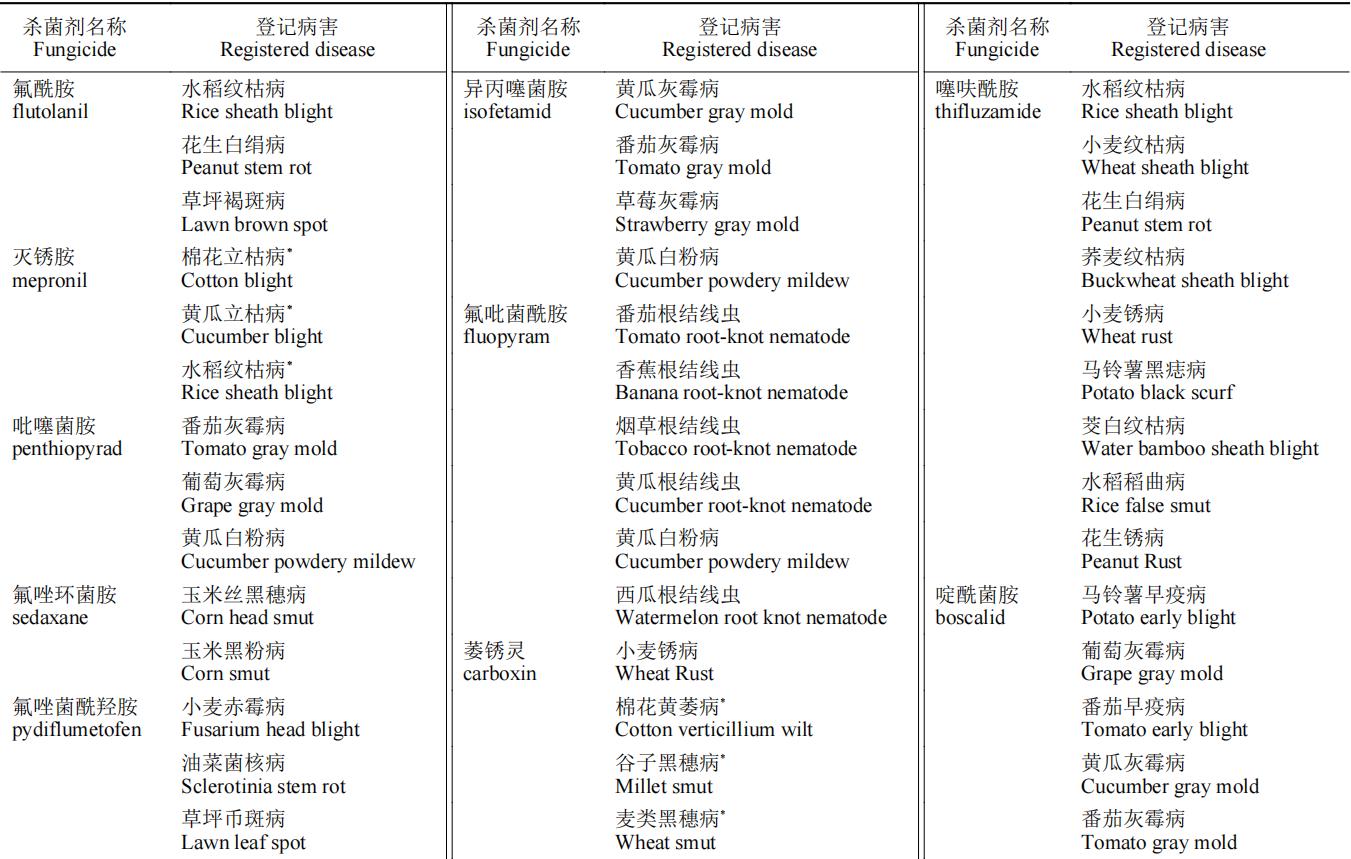
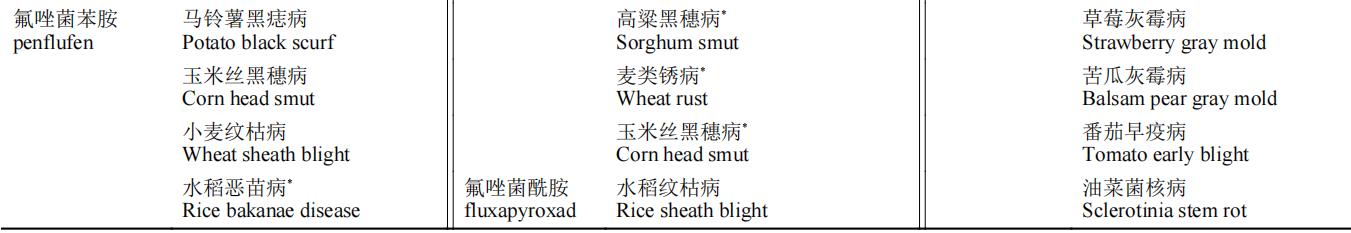
注:“*”表示曾被登记防治,现今已过登记有效期。
SDHIs杀菌剂主要登记用于担子菌、子囊菌和半知菌引起的植物病害的化学防治。由于SDHIs杀菌剂作用位点单一,抗性风险高,在产品登记上与其他产品复配成为了较好选择。从目前上市的品种看,SDHIs杀菌剂与主流的Qo位点呼吸抑制剂类和麦角甾醇生物合成抑制剂类杀菌剂组合大大开拓了应用范围和防治谱。
3 SDHIs杀菌剂抗性发生现状
SDHIs杀菌剂的固有抗性风险被FRAC归类为中至高等。最早的SDHIs杀菌剂抗性报道至少可追溯至1975年,Gunatilleke等在实验室条件下通过诱导试验获得了萎锈灵(carboxin)抗性的构巢曲霉菌株(Aspergillus nidulans)。21世纪以来,杀菌剂的抗性问题逐渐引起人们的重视,与此同时SDHIs杀菌剂的应用范围也愈发广泛,使用年限越来越长,该类药剂的田间抗性问题也逐渐加重。目前,SDHIs杀菌剂的抗性问题已经发生在多种病原菌上,涉及多个国家。在美国已检测到开心果腐烂病菌(Alternaria alternata)抗啶酰菌胺(boscalid)菌株,且对萎锈灵(carboxin)表现出正交互抗性;番茄早疫病菌(Alternaria solani)抗SDHIs菌株,马铃薯早疫病菌(A. solani)抗啶酰菌胺(boscalid)和吡噻菌胺(penthiopyrad)菌株等。在巴西,在小麦麦瘟病菌(Pyricularia oryzae)中检测到氟唑菌酰胺(fluxapyroxad)的抗性菌株,菊花白锈病菌(Puccinia horiana)种群中也已经检测到SDHIs杀菌剂抗性。在欧洲SDHIs杀菌剂抗性也已经在多种病原菌中发生,如小麦壳针孢菌(Zymoseptoria tritici),苹果黑星病菌(Venturia inaequalis),马铃薯早疫病菌(A. solani),百合叶枯病菌(Botrytis elliptica),油菜菌核病菌(Sclerotinia sclerotiorum)等。在中国,小麦赤霉病菌(Fusarium graminearum)、番茄灰霉病菌(Botrytis cinerea)、桃腐烂病菌(A. alternata)、黄瓜靶斑病菌(Corynespora cassiicola)等病原菌中均已经出现SDHIs杀菌剂抗性种群。随着SDHIs杀菌剂使用年限和范围的增加,抗药性问题呈现加重趋势,延缓和治理SDHIs杀菌剂的抗性已经成为一个迫切需要解决的重要科学问题。
4 SDHIs杀菌剂抗性分子机制
4.1 靶标基因点突变
植物病原真菌药靶基因的遗传分化决定了杀菌剂的选择性,不同植物病原真菌药靶基因编码的氨基酸数量也存在一定差异。敏感的植物病原菌药靶基因发生特异性突变可以导致对杀菌剂产生不同水平的抗性。Mair等建议,将所有的同类型杀菌剂抗性点突变进行规范化,统一命名,为杀菌剂靶标蛋白突变提出了一个统一的氨基酸标记系统,即不论实际位置如何,同源氨基酸位点都被赋予相同的数字,便于人们理解抗药性位点。线粒体复合物Ⅱ通常由SDHA、SDHB、SDHC和SDHD等4个亚基组成,在已有的报道中,还未见SDHA亚基上出现抗药性突变,已知主要的抗药性突变位点集中在SDHB、SDHC、SDHD等3个亚基上。
随着SDHIs杀菌剂使用年限的增长,抗性报道也逐步增多。根据近年来的相关抗性机制研究报道和FRAC的统计结果,植物病原菌的抗药性突变基因型归纳于表3。
表3 植物病原菌与SDHIs杀菌剂抗性相关的点突变基因型
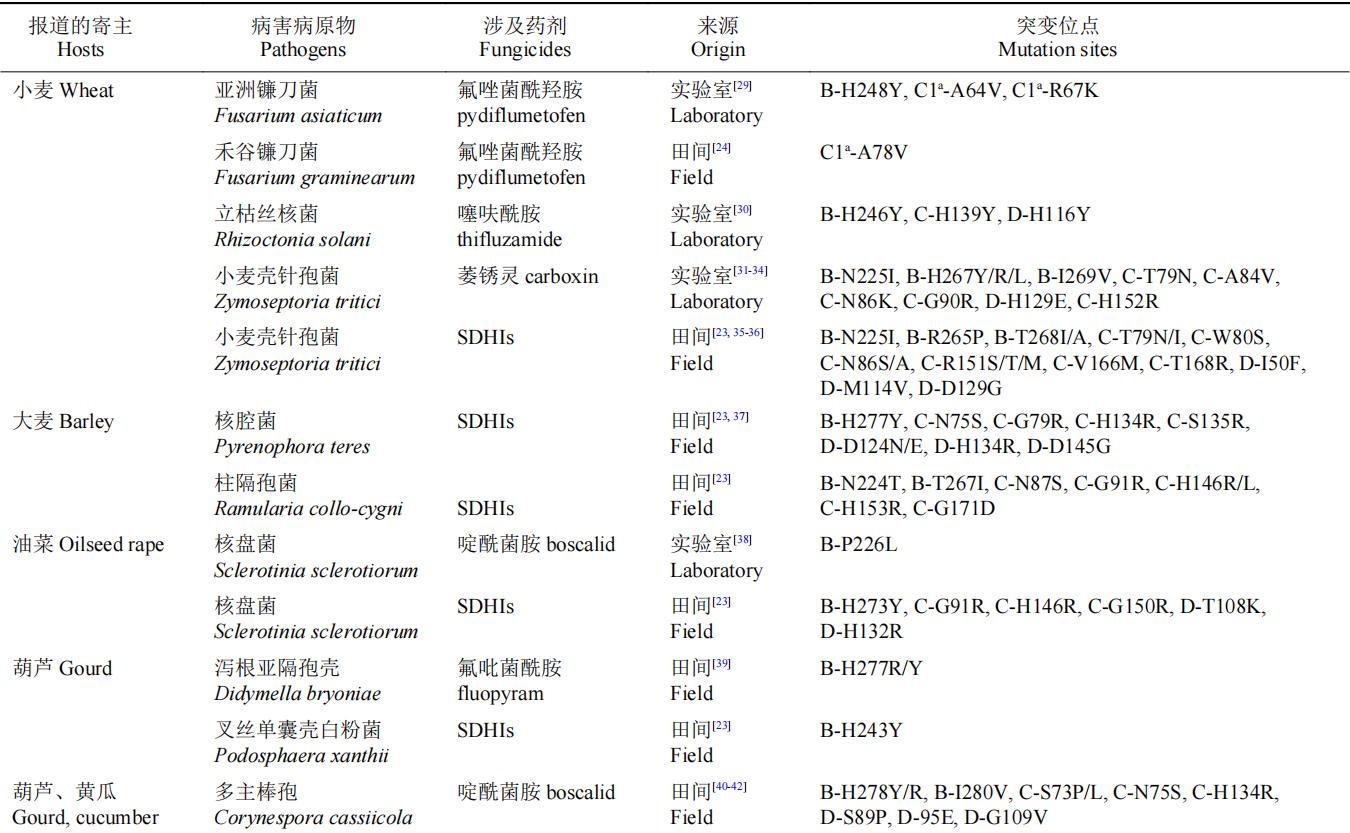
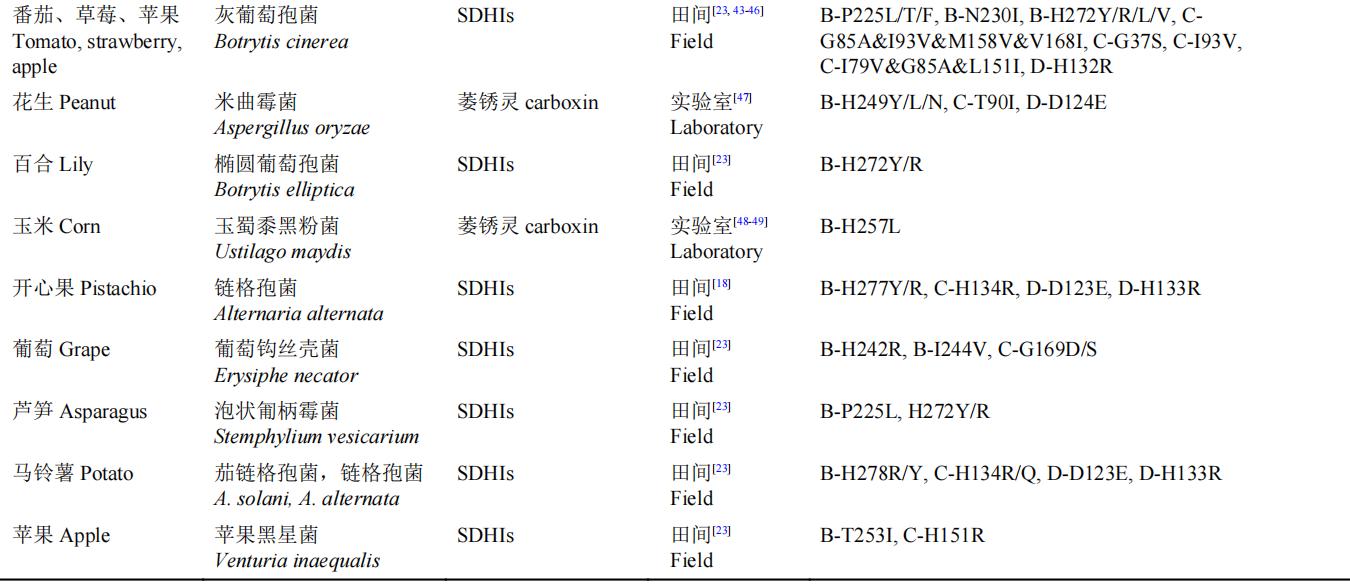
a:由于小麦赤霉病菌种存在两个SDHC亚基基因,以C1、C2区分。
近年来,FRAC联合巴斯夫、拜耳、科迪华、先正达、富美实、安道麦、住友集团等企业在美国、巴西、欧洲等地区进行了多种病原的抗性监测工作,并逐年在FRAC官网上发布结果。在SDHB、SDHC、SDHD亚基上的抗药性突变形式有很多,甚至同一物种中存在各种不同形式的抗药性点突变基因型。在以上突变中SDHB亚基的突变在257、267或272位由组氨酸突变为酪氨酸/异亮氨酸/精氨酸,225位脯氨酸突变为异亮氨酸,在多物种中相对比较保守。但并非所有的田间突变都能引起药敏性的变化,例如,小麦壳针孢菌(Z. tritici)田间抗药性群体的突变B-C266G、C-N33T、C-N34T、C-L184W已被验证与SDHIs杀菌剂药敏性无关。本研究团队最新研究发现,核盘菌SDHB-A11V突变并非SDHIs杀菌剂抗性突变位点,而SDHB-P226L能引起SDHIs杀菌剂的中等水平抗性。
在抗药性进化过程中,某些突变会伴随一些生物适合度的下降,如生长速率、致病力等。此外,由于一些真菌的多核现象,杂合突变体的抗药性一直无法准确鉴定。我们在开展核盘菌(多核丝状真菌)对SDHIs杀菌剂抗性的相关研究中,通过人工点突变技术构建了SDHB-P226L的点突变载体,并通过遗传转化技术,同源置换野生菌株中的SDHB基因,获得的转化子多为杂合子,生物适合度低,进一步对杂合子进行药剂驯化,获得了点突变纯合子菌株,抗药性水平与生物适合度均显著提高。刘西莉研究团队在立枯丝核菌(多核丝状真菌)的抗药性研究中,对杂合体抗药性突变菌株进行药剂驯化,完成了杂合子纯合化的遗传变异,演化了真菌的抗药性进化过程。Bart研究团队建立了一套系统的剂量依赖型杀菌剂抗性诱导技术,用于评估SDHIs杀菌剂的抗性风险。
4.2 SDHIs杀菌剂的非靶标抗性
Yamashita等率先在小麦壳针孢菌上报道了SDHIs杀菌剂的非靶标抗性。在欧洲采集的田间小麦壳针孢病原群体中(Z. tritici)发现了可以稳定遗传的抗药性菌株,但在其SDHA、SDHB、SDHC、SDHD亚基上并未检测到抗药性突变位点。在交互抗性测定中发现,此种抗性仅仅存在于包括氟吡菌酰胺(fluopyram)和异丙噻菌胺(isofetamid)在内的部分长杂环酰胺类SDHIs杀菌剂,而对联苯吡菌胺(bixafen)和氟唑菌酰胺(fluxapyroxad)等其他酰胺类SDHIs杀菌剂无正交互抗性。非靶标抗药性菌株和相同作用靶标的杀菌剂之间无交互抗性的发现,引起了人们对SDHIs杀菌剂抗性与药剂靶标的深入思考和探究。
通常情况下,靶标基因的突变和过表达是其对杀菌剂产生抗性的主要原因,但目前尚未有琥珀酸脱氢酶相关亚基基因过表达而引起抗药性的相关研究。然而,Mae等发现在小麦壳针孢菌(Z. tritici)中,MFS1(major facilitator superfamily)启动子区的转座子插入引起了对SDHIs杀菌剂的抗性;Sang等在对氟吡菌酰胺(fluopyram)不敏感的大豆猝倒病菌(F. virguliforme)菌株中,发现MFS转运体基因过量表达,同时,在酵母系统中,通过异源表达验证了这种耐药性。这种耐药性产生的原因为外源性物质解毒相关基因过表达引起细胞对药剂的外排作用增强从而减弱杀菌剂的毒力。
4.3 长杂环酰胺类SDHIs与普通SDHIs杀菌剂的抗性差异
我们研究发现从黄瓜上分离的多主棒孢(C. cassiicola)发生SDHB-H278R点突变导致对啶酰菌胺(boscalid)产生抗药性,但对SDHI杀菌剂氟吡菌酰胺(fluopyram,属于长杂环酰胺亚类)不存在交互抗性。张晓珂等发现,在灰霉病菌中,氟吡菌酰胺(fluopyram,属于长杂环酰胺亚类)与啶酰菌胺(boscalid)之间无交互抗性。Ishii等发现,在多主棒孢(C. cassiicola)和瓜类白粉病菌(Podosphaera xanthii)中,啶酰菌胺(boscalid,非杂环酰胺亚类)和吡噻菌胺(penthiopyrad,非长杂环酰胺亚类)抗性菌株对氟吡菌酰胺(fluopyram,属于长杂环酰胺亚类)未表现出交互抗性。这些研究结果表明,病原菌对长杂环酰胺亚类SDHIs与普通SDHIs的抗性机制存在差异。
Steinhauer等以“产生非靶标抗药性的小麦壳针孢菌”为研究材料,探究了同类SDHIs杀菌剂无交互抗性的原因。通过正向遗传学技术分析得到小麦壳针孢菌(Z. tritici)部分种群中存在多个SDHC同源基因;并结合反向遗传学技术,揭示了小麦壳针孢菌(Z. tritici)非靶标抗药性产生的原因是由于SDHC3基因的存在,且抗性水平和SDHC3的表达水平及选择性剪切效率的差异有关;同时,SDHC3启动子区的转座子插入也介导了SHA-SDHI杀菌剂的抗性。有趣的是,SDHC3的存在仅对SHA-SDHI杀菌剂表现抗性,对其他非SHA-SDHI杀菌剂并未表现出抗性,点突变策略和分子对接也进一步证实了SDHC3为SHA-SDHI杀菌剂的药剂靶标。这一发现从分子角度揭示了“药-靶”互作机理,丰富了当前的SDHIs杀菌剂的抗性机制研究。
4.4 SDHC亚基遗传分化对SDHIs杀菌剂药敏性的调控研究
有趣的是,在不同的植物病原真菌中,琥珀酸脱氢酶的C亚基可遗传分化为多个亚型,且调控着对SDHIs杀菌剂的药敏性。邵文勇等发现在灰霉病菌(B. cinerea)中,存在5种不同的SDHC分化亚型,不同亚型的灰霉病菌对SDHIs杀菌剂的药敏性存在一定的差异。灰霉病菌SDHC不同亚型的遗传分化可能是在SDHIs杀菌剂的选择压下导致的。然而核盘菌(S. sclerotiorum)中,SDHC亚型的遗传分化似乎与SDHIs杀菌剂药敏性并无联系,本团队在核盘菌中发现SDHC亚基存在两种亚型,两者存在11个氨基酸的差异,在田间病原群体的比例约为7∶3,但核盘菌SDHC亚基遗传分化的两种亚型对SDHIs杀菌剂药敏性并无显著差异。
Steinhauer等发现小麦壳针孢菌(Z.tritici)SDHC遗传分化为3个亚基(SDHC1、SDHC2、SDHC3),并证实SDHC3是SHA-SDHI杀菌剂的药剂靶标。李美霞等发现在小麦赤霉病菌中存在两个不同SDHC亚基(SDHC1、SDHC2),共同调控着SDHIs杀菌剂的药敏性。当SDHC1敲除时,小麦赤霉病菌对SDHIs杀菌剂的药敏性显著增加,表现出超敏感,而当SDHC2敲除时,小麦赤霉病菌对SDHIs杀菌剂的药敏性显著降低,表现出抗药性。此种调控机制,正如苯并咪唑类杀菌剂药靶基因β-tubulin在小麦赤霉病菌中的遗传分化,β1-tubulin负调控对微管蛋白抑制剂的抗药性,而β2-tubulin正调控对微管蛋白抑制剂的抗药性。这种药靶基因遗传分化对药敏性表现出双向调控的现象,暗示了SDHC1、SDHC2可能竞争性地参与或调控着琥珀酸脱氢酶复合体的结构形成,但其调控方式仍需我们通过蛋白质互作与结构生物学技术进一步验证。
5 SDHIs杀菌剂抗性检测/监测方法
随着单一作用位点的现代选择性杀菌剂的飞速发展,抗药性问题已成为必然。因此,开展抗药性检测/监测研究,不仅能实现植物病原菌的抗药性流行预警,而且可指导植物病害防控的精准选药、科学施药,保障我国的农药减量与农业生产安全。抗药性检测/监测技术是随着现代选择性杀菌剂的问世不断发展起来的,可以分为以下3个阶段:
第一阶段是传统的生物测定法,通过药剂对菌丝生长或孢子萌发抑制的差异来判定是否具有抗药性,通过大量的抗性监测数据和药敏性测定结果,优化出区分剂量,来筛选抗性菌株,可减少工作量;如:桃黑斑病菌(A. alternata)等的SDHIs杀菌剂的抗性检测。该方法需要经过菌株的分离、纯化、培养,在含药平板上进行抗药性检测,由于工作量大、周期长、效率低、检测成本高,以致检测的菌株数量有限,往往在检测到抗性菌株时,病原菌群体中抗药性菌株的比例已经达到1%以上,在短期内即可造成抗药性病害流行,造成突发性危害。
第二阶段的检测技术,主要是基于药靶基因突变后与野生菌株的核酸序列差异而开发的分子检测方法。这一阶段的检测技术往往以PCR为基础,最为有效、直接、准确的方式是通过扩增靶基因序列并测序,分析突变位点。基于序列差异,利用限制性内切酶对扩增后的多态性核酸序列进行裂解,通过电泳分析。这种方法相对于测序的成本大大降低,被广泛用来进行抗药性突变检测。其中,限制性片段长度多态性(RFLP,restriction fragment length polymorphism)方法被用于检测开心果晚疫病(A. alternata)、多寄主灰霉病(B. cinerea)对SDHIs杀菌剂抗药性的点突变。裂解扩增多态性序列(CAPS,cleaved amplified polymorphic sequence)被用于Clarireedia属SDHIs杀菌剂抗性的检测。等位基因PCR(AS-PCR,allele specific PCR)被用于A. alternata种SDHIs的抗性检测。一种基于PCR的高分辨率溶解曲线法(HRM,high-resolution melting)用来检测灰霉病菌(B.cinerea)SDHB-H272R/Y序列变化。这一阶段的抗药性分子检测技术,能够在几小时内鉴定或诊断菌株的抗药性,高通量定量PCR技术能够在病原群体中检出万分之一至十万分之一的抗药性基因频率,实现了抗药性早期预警,解决了抗药性病害突发和防控措手不及的问题。但是,这阶段发展起来的分子检测技术,需要昂贵的仪器和高级技术人员才能完成,仍然限制了这些技术的推广应用。
第三阶段是基于环介导恒温扩增技术(Loop-mediated isothermal amplification,LAMP),本研究团队创造性地研究了基因扩增引物的碱基错配技术,率先发明了可诊断药靶基因单碱基突变的LAMP抗药性高通量快速检测技术,借助LAMP技术,在检测水平上实现了质的飞跃。2014年,本研究团队率先将LAMP技术结合到植物病原菌抗药性检测方面,开发了一系列配套技术,大大革新了病原菌抗药性检测技术,此后,针对SDHIs杀菌剂抗性位点的LAMP检测技术也被开发。LAMP检测技术无需昂贵的仪器设备和繁琐的试验操作过程,检测灵敏度与检测效率得到进一步提高,相应的检测技术已开发为快速检测试剂盒,解决了检测的时空限制问题,检测条件极度简化,田间地头即可快速检测。一台水浴锅或恒温箱反应一小时,即可通过肉眼观察样品颜色差异来判断结果,简便快捷,非常适合田间快速诊断,但是仍然存在着只能对抗药性进行定性分析的缺陷。目前,本研究团队正在研发新一代LAMP抗药性定量快速分子检测技术,该技术研发成功有望将抗药性检测效率再提高100倍以上,检测成本降低至1%,未来的定量快速分子检测技术十分令人期待。
6 SDHIs杀菌剂抗性风险与治理
FRAC将SDHIs杀菌剂的固有抗性风险列为中至高等抗性风险。长期单一使用SDHIs杀菌剂,植物病原真菌对这类杀菌剂极易产生抗药性。尽管病原菌的SDHIs杀菌剂靶标基因存在低、中、高水平抗药性基因型,但同一种SDHIs杀菌剂的连续使用和加量使用,会杀死或抑制敏感和低抗水平的病原菌,加速形成中、高水平抗药性群体,使SDHIs杀菌剂失效。因此,延缓或阻止抗药性群体发展是抗药性治理的重要目标。
由于药靶基因在不同的病原菌中存在分化,并存在不同的保守型,因此,在杀菌剂进入田间防治前,应建立敏感性基线和开展抗性风险评估,为科学用药和抗性监测提供科学依据。目前已有报道中,SDHIs杀菌剂抗性菌株的抗性水平主要为中低水平抗性,相较敏感群体,EC50值倍数在5~30之间,此种抗性水平在药剂选择下,有利于抗性群体的发展。
病原菌对杀菌剂发生抗性变异后的适合度是影响抗性群体发展的重要因素。桃腐烂病菌(A. alternata)SDHD-D123E点突变的抗性菌株,产孢量下降,在氧化应激压力下生长速率降低,说明在没有药剂选择压下,抗药性病菌在群体中的比例会下降。然而,马铃薯叶枯病菌(A. alternata和A. solani)携带SDHB-H277Y/R、SDHD-D123E点突变菌株无明显适合度变化,草莓灰霉病菌(B. cinerea)携带SDHB-H272R/Y/L,SDHB-P225F,SDHB-N230I点突变的菌株也无明显适合度变化。大多数研究表明,SDHIs杀菌剂抗性病原菌的适合度与野生敏感群体相比,并无明显下降,说明在药剂选择下,抗药性病原群体会发展较快。在杀菌剂进入田间应用以后,实时动态监测/检测田间病原菌的抗药性发生动态,对于抗药性早期预警、储备抗药性治理技术、评价抗药性治理策略及技术的有效性等具有重要意义。
抗药性病害流行不仅取决于抗药性菌株在群体中的比例,更取决于抗药性病原菌的绝对数量。当病原菌群体在自然界的数量巨大时,即使抗药性病菌未成为优势种群,在药剂防治时剩下的抗药性病菌也足以引起病害流行。因此,能够降低药剂选择压,如利用抗病品种、生物防治等病害综合防治策略均有利于延缓抗药性群体发展。Samaras等发现,生防菌(Bacillus amyloliquefaciens)在与氟吡菌酰胺的交替使用中对灰霉病表现出较高的抑制作用,并降低了SDHIs类杀菌剂的抗性频率。病害发生严重度直接关系到病原群体的发展,凡是有利于降低病害压力的农艺措施,如田园卫生及通风透光等栽培措施均是抗药性治理策略与技术的重要组成部分。
SDHIs杀菌剂种类多,结构差异大,尽管FRAC给予了这些杀菌剂相同的抗性分类编码(#7),属于同一交互抗性组,但是目前已经发现一些病原菌对长杂环酰胺类SDHIs杀菌剂和普通SDHIs杀菌剂并无交互抗性。因此作者认为,在明确所防治的病原群体对SDHIs杀菌剂之间交互抗性条件下,或可考虑混用或轮用不同亚类的SDHIs杀菌剂或以不同类型SDHIs杀菌剂来缓解SDHIs杀菌剂抗性。
SDHIs杀菌剂推荐与其它作用机制的药剂(Qo位点呼吸抑制剂类杀菌剂、麦角甾醇抑制剂类杀菌剂)以复配的方式使用时,不仅应该遵循增效原则和无药害原则,而且必须考虑是否有利于抗药性治理。不同作用方式的杀菌剂如果存在相同的抗药性机制也不应该混合使用,如相同的增加药剂外排抗性机制、相同化学基团的降解与解毒机制等。值得注意的是,那些抗性机制和作用方式不同的两种高抗性风险杀菌剂也应谨慎混合使用,防止在短时间内病原菌对两种杀菌剂均产生抗药性,枯竭有效的杀菌剂资源,如SDHIs杀菌剂与QoIs杀菌剂混剂防治蔬菜灰霉病,很快使灰霉对这两种类型的杀菌剂产生了高水平抗性群体。
研发作用机制新颖、没有交互抗性的新型杀菌剂是治理抗药性的根本措施。针对药靶变异的生物学信息设计和创制反抗性杀菌剂,对于治理重要杀菌剂分子靶标抗性具有重要价值。然而,利用现代生物学技术挖掘和发现杀菌剂新的分子靶标、发现调控药-靶互作的关键蛋白、解析和发现靶标蛋白新的药敏性结构域等,是靶向新型杀菌剂和靶向增效剂创制的理论基础。
7 展望
当前,SDHIs杀菌剂在全球杀菌剂市场中仍处于上升期,SDHIs杀菌剂仍有广阔的市场需求,因此开展SDHIs杀菌剂的抗性研究,对于延缓其抗药性发展速度,开发新型高活性的SDHIs杀菌剂仍具有重要意义。同时当前的SDHIs杀菌剂与植物病原菌药剂靶标之间的研究结果也加深了人们对药靶互作的理解,丰富了SDHIs杀菌剂的作用机制研究,为SDHIs杀菌剂的反抗性药剂或者难以产生抗性的新药剂研发提供了数据基础。自然界天然的与SDHIs杀菌剂结构类似的活性物质,与当前的酰胺类杀菌剂具有不同化学结构,这也启发了科学家开展新型SDHIs杀菌剂的研发,同时也从侧面印证了SDHIs杀菌剂的环境安全性。此外,通过修饰和改造现有杀菌剂,研发高选择性、高活性、高安全性、低风险的SDHIs杀菌剂仍在进行。各种抗药性检测技术的发展也为SDHIs杀菌剂的抗药性监测提供了技术保障。可以预见,SDHIs杀菌剂仍将在未来的农药市场中发挥中流砥柱的作用。
(1)本网旨在传播信息,促进交流,多方面了解农药发展动态,但不构成任何投资建议。
(2)所有文章仅代表作者观点,不代表本网立场。
(3)“信息来源:江苏省农药协会 农药资讯网”为原创文章,转载时请注明来源和作者。
(4)本网转载文章及图片的版权属于原作者,若有侵权,请联系删除。